|
|
ВОЛНОВАЯ ГЕНЕТИКА
Гаряев Петр Петрович
доктор биологических наук, академик РАМТН (Российская Академия медико-технических наук) и РАЕН (Россиская Акдемия Естественных Наук), президент Института квантовой генетики, научный директор Wave Genetics Inc. (Канада, Москва)
E-mail: gariaev@mail.ru
СОДЕРЖАНИЕ
· Генетические структуры как источник и приемник голографической информации
· Перспективы волновой генетики
· Волновые репликативные отображения ДНК и ее ближайшего окружения
· Другие материалы а также обсуждение темы Волновая Генетика на форуме
Генетические
структуры как источник и приемник голографической информации
Прангишвили И.В., Гаряев П.П., Тертышный Г.Г., Леонова-Гаряева Е.А., Мологин А.В.
Живое и Неживое имеют общие
характеристики информационно-голографического характера. Это связано с
фундаментальной способностью когерентных физических полей интерферировать и,
тем самым, при наличии регистрирующей среды, “записывать” собственные
характеристики. Существенно, что в этом процессе не играет роли
какие структуры, Живые или Неживые, порождают эти поля.
Развивая эти идеи, опубликованную нами ранее [1, 2], в настоящей работе мы уточняем нашу гипотетическую модель эндогенного биоволнового (голографического) управления развитием многоклеточных организмов из эмбрионального состояния во взрослое. Кроме того, в последние годы выясняется, что к числу фундаментальных явлений природы относится так называемый “фоновый принцип” [3], непосредственно связанный с биоголографической работой генетического аппарата [1, 2]. Суть фонового принципа заключается в том, что известное соотношение шум/сигнал трактуется так, что уровень сигнала можно повысить не за счет подавления шума (фона), как это делается обычно, а наоборот, фон служит источником сигнала и средством повышения его уровня. Этот принцип универсален и работает во всех областях человеческого знания, в общем отображая категории Части и Целого, в том плане, что полное знание о части невозможно без учета свойств целого.
Например, в лингвистике фоновый
принцип хорошо виден в случае омонимий, когда слова-омонимы (‘лук’ ‘рейд’
‘коса’ и т.д.) не могут иметь точную семантику вне целого предложения, то есть
контекста (фона). В генетической лингвистике применение
фонового принципа позволило нам решить [2] парадокс Вобл-гипотезы Ф.Крика, в
соответствии с которой в генетическом коде существует задача снятия
омонимических неоднозначностей знаковых дублетов кодонов. Эту проблему
живая клетка, точнее ее белок-синтезирующий
аппарат, снимает именно на основе фонового принципа за счет контекстных
ориентаций рибосомы в геометрически одномерном смысловом пространстве
информационных РНК. В свою очередь, такие контекстно-фоновые механизмы
“логической” ассоциативной работы рибосомы невозможны без интегрально-волнового
восприятия целого генетического текста информационных и, возможно, преинформационных РНК.
Другой вектор приложения логики
фонового принципа заключается в том, что в радиоволновом диапазоне можно
обнаруживать и опознавать макрообъекты-“невидимки”, изготовленные по технологии
“Стелс”. Фоновый принцип делает вполне реальной разрабатку микроскопии (в дополнении к фазово-контрасной)
прозрачных для света живых и динамичных биоструктур
без повреждающей предварительной окраски. Можно наблюдать, например, сложную
знаковую динамику хромосом в живых клетках и связать ее с волновыми
управляющими функциями генетического аппарата, а это до сих пор было недоступно
для исследователей.
Известно, что хромосомный
континуум многоклеточных организмов является высокоорганизованной системой
генерации эндогенных когерентных электромагнитных полей широкого спектра от 250
нм до 800 нм, далее ИК- и СВЧ-диапазоны
[1, 2]. Высшим биосистемам свойственна также знаковая акустика генетических
структур, которая может иметь солитонный и
когерентный характер. Эти информационно-волновые процессы распространяются и на
белковую, и на водную составляющие биосистем [1, 2].
Поэтому практически любая точка в пространстве организмов является источником
физических полей, в том числе генознаковых. Все
организмы земли взаимодействуют с внешними естественными и искусственными
электромагнитными и акустическими полями, некоторые из которых могут быть
когерентными, например, поля от таких удаленных источников, как звезды.
Имея в виду эти соображения,
применим принцип Гюйгенса к биосистемам и будем считать, что каждая точка живой
клетки является источником элементарной электромагнитной и/или акустической
волны. Комплексная амплитуда волны в каждой такой точке равна сумме комплексных
амплитуд отдельных элементарных волн, излучаемых каждой точкой биосистемы
(далее речь идет только об электромагнитных волнах). Поскольку нас, прежде
всего, интересуют генетические волновые процессы, будем рассматривать в
качестве источников элементарных волн управления жидкокристаллические фазы ДНК
в составе хромосомного континуума. Характерным для таких фаз как in vitro
так и in vivo является их
сложная высокоорганизованная надмолекулярная слоистая структура, состоящая из холестерических сверхплотных укладкок
спиралей ДНК [4]. Такие слои ДНК можно рассматривать как аналоги транспарантов
в голографии.
Физико-математическая модель процесса излучения волны молекулами ДНК записывается следующим образом. Допустим, что в некоторой плоскости координат x,y размещен ограниченный элементарный плоский слой жидкокристаллической ДНК, слой, который образовал излучательный континуум. Тогда элементарный фронт волны, исходящей из произвольной точки x на слое ДНК, представим в виде
![]() где
где ![]() -
расстояние между точкой
-
расстояние между точкой![]() и
точкой
и
точкой![]() , в которой определяется поле.
, в которой определяется поле.
Суммарная амплитуда элементарных
волн в точке![]() запишется
в виде
запишется
в виде
![]() (1) .
(1) .
При интегрировании в бесконечных пределах соотношение (1) с точностью до постоянного члена перед интегралом является формулой дифракции. Ввиду того, что расстояние rs есть иррациональная функция разности координат, ограничимся небольшой областью в окрестности оси z и применим параболическое приближение
![]() , где Zs - продольное расстояние между объектной точкой на
участке плоской структуры ДНК и точкой, в которой определяем уровень светового
электромагнитного напряжения.
, где Zs - продольное расстояние между объектной точкой на
участке плоской структуры ДНК и точкой, в которой определяем уровень светового
электромагнитного напряжения.
Если бы в эту точку мы могли поместить светочувствительную среду, то в ней возможна регистрация голограммы зондируемого участка биосистемы при наличии референтной электромагнитной волны [5]. Такая ситуация возможна, если считать, что светочувствительными средами являются сами жидкие кристаллы ДНК в составе хромосом и/или структурированная вода биосистемы, точнее ее электролиты, составляющие около 70% ее объема. Это достаточно реалистично, поскольку голографическая память генома получила экспериментальные подтверждения, а коротко живущие голограммы на водных растворах известны в техническом исполнении [1]. Однако, можно обойтись и без этого допущения, если в применении к геному биосистем дополнительно использовать понятие условных голограмм (квазиголограмм).
Отличие голограммы от квазиголограммы состоит в способе их формирования (записи). Для регистрации обоих типов голограмм требуются опорные (референтные) волны. Однако, для квазиголограмм референтные волны могут быть фрагментами объектных волн и выступать как квазисферические за счет дифракции на отдельных микроструктурах биосистемы. По этой причине формируются многоопорные референтные пучки для формирования градиентов поля квазиголограммы. Такого рода квазиголограммы будем называть волновыми (в-квазиголограммы).
Квазиголограммы ДНК могут формироваться и другим путем, который зависит от последовательностей нуклеотидов, определяющих характер жидкокристаллических упаковок хромосом и, следовательно, структуру (информацию) квазиголограмм [1]. Их мы будем называть последовательность-зависимыми (п-квазиголограммами). Однако, в чистом виде ‘в’- и ‘п’- типы вряд ли существуют. Скорее всего, формируются смешанные типы. Есть основания думать, что в биосистемах образуются и квазиголограммы, и голограммы [1, 2]. Поэтому, далее в статье употребление этих терминов по биологическому смыслу (конечному биорезультату) семантически тождественно.
Ключевую роль в волновом управлении формирования таких голографических решеток могут играть факторы изменения градиентов напряженности физических полей, пронизывающих гетерогенное пространство организмов и квантованно (по углу регистрации-считывания, по длине волны и по субстратам записи) записывающих - воспроизводящих его динамичную структуру. Эти градиенты зависят от конкретных биоструктур и от голограмм - квазиголограмм, образуемых ими. Именно на них дифрагируют фотонные и другие поля с образованием знаково-непрерывного континуума градиентов этих полей. Градиенты в данном случае выступают как носители биознаков пространственной разметки биосистем, а совокупность градиентов образует знаковый континуум построения многоклеточного организма. Важным моментом волновой организации биосистем является фактор безынерционной радиосвязи по поляризационному каналу между ядрами клеток биосистемы для их синхронизации, введения в когерентное состояние “знания” друг о друге. Такие радиоволновые излучения могут индуцироваться лазерными излучениями хромосом по типу обнаруженного явления конверсии фотонов в радиоволны [2, 6, 7]. В тесной связи с этим ключевой (в биологическом смысле) является предполагаемая нами фрактально масштабированная когерентность (связь) по каналу поляризации фотонов с радиоволнами (Ф® Р гипотеза) на уровнях хромосомного континуума и других управляющих организмом биоструктур, таких как ферментные ансамбли, цитоскелет, внеклеточный матрикс и т.д. В рамках Ф® Р гипотезы снимается проблема радиоволновой “прозрачности” организмов и, соответственно, биологической активности радиоволн, знаково модулированных по поляризации. Действительно, существуют противоречивые данные, связанные с “прозрачностью”, т.е. отсутствием выраженного действия хаотически поляризованных радиоволн кило- и мегагерцового диапазонов на живые организмы. Можно полагать, что эти диапазоны будут так или иначе воздействовать на биосистемы, если они имеют знаконесущие поляризационные модуляции. Это означает, что они будут вызывть биологический (метаболический) ответ при наличии поляризационных резонансов с радиоизлучающими и радиопринимающими командными биоструктурами, например, с ДНК, РНК и белками.
Подставив выражение (2) в (1), мы получим формулу дифракции Френеля. Дифрагирующая объектная волна “записывает” информацию о геометрии (форме) биоучастка, на котором происходит акт дифракции. В простейшем варианте это можно представить в виде ряда событий. Волна, излученная слоем хромосомной ДНК, выходя за пределы клеточного ядра, дифрагирует на встречающихся клеточно-тканевых структурах, то есть модулируется по амплитуде, фазе, поляризации, частоте и тем самым приобретает информацию о строении этих структур. Это фактор автоконтроля биосистемы за своей пространственной структурой [1, 2]. Для регистрации в форме голограммы полученной информации нужна референтная волна, которую представим как сферическую и запишем ее также в параболическом приближении
![]() Здесь
Здесь ![]() -
амплитуда, а
-
амплитуда, а ![]() -
местоположение референтной волны, которое можно
выразить через координаты
-
местоположение референтной волны, которое можно
выразить через координаты ![]() и
и![]()
![]() (4) .
(4) .
В результате сложения комплексных амплитуд объектной и референтной волн получим выражения для интенсивности, равной квадрату абсолютной величины комплексной амплитуды результирующего поля
![]() (5) .
(5) .
Раскрывая квадрат от модуля комплексной амплитуды суммарного поля, получим
![]() (6)
(6)
![]()
![]()
Можно полагать, что, следуя закону распределения интенсивности поля или его градиенту, биосистема будет использовать эти градиенты как некоторые (из многих) векторы построения своей клеточно-голографической структуры именно в тех местах, которые соответствуют амплитудному пропусканию голограммы в соответствии с условным перераспределением интенсивности и связанным с ним ростом и развитием биоструктур, взаимодействующих с фотонами. Возникает понятие “условного потемнения”, которое мы понимаем как интерференционный результат сложения когерентного излучения по амплитуде, фазе, поляризации и частоте. Эти изменения модуляции интенсивности поля происходят вследствие сгущений и разряжений оптической плотности при динамике клеточно-тканевых структур в ходе развития и жизнедеятельности биосистем. Законы этих сгущений и разряжений и перемещений, т.е. картины и сценарии интенсивности организменных полей, в том числе их оптическая активность и спектральный состав, являются содержанием цитодифференцировок и морфогенеза организмов в волновом представлении. Заметим, что под частотной (спектральной) модуляцией фотонов, проходящей на биоструктурах, надо понимать не только и не столько известное комбинационное рамановское преобразование частот в видимом и инфракрасном интервалах длин волн. Мы допускаем, что в биосистемах может иметь место аналог феномена, обнаруженный нами ранее [2, 6, 7]. Он, в частности, проявляется в трансформации когерентного света, в определенных условиях взаимодействующего с геноструктурами и белками, в биологически активные радиоволны кило- и мегагерцового диапазонов. При этом, как уже упоминалось, существенна корреляционная зависимость поляризации фотонов и поляризации рожденных из них радиоквантов как потенциальный биоинформационный канал. Не исключено, что этот канал является модификацией открытого недавно эффекта квантовой телепортации связанных по поляризации фотонов, предсказанного в 1935 г. как эффект квантовой нелокальности (квантовой телепортации) Эйнштейна - Подольского - Розена [9]. Возможно, этот канал обеспечивает адекватное восприятие и использование биосистемами внутренней и внешней волновой среды их обитания.
Для правильного понимания предлагаемой голографической модели организации биосистем, надо учитывать, что квази - спонтанная динамика биоструктур in vitro - in vivo, по крайней мере, для таких ключевых как ДНК, рибосомы и коллаген, носит нелинейный квантованный характер и реализуется в рамках явления возврата Ферми-Паста-Улама (ФПУ), т.е. является солитонным процессом с памятью на начальные условия возбуждения колебательных мод. Указанные биоструктуры, возможно, имеют квантованный спектральный состав акустических колебаний с меняющимися, но периодически повторяющимися некоторыми модами [1]. Здесь физика колебательных процессов обеспечивает фундаментальный атрибут биологической памяти генетических молекул, а именно их способность к волновому “самопрочитыванию” генетических “текстов” и голограмм с возвратами ФПУ к уже “прочитанному” и/или “просмотренному” через высокоорганизованные и квантованные вещественно-волновые состояния, как особого рода “памяти” хромосом. Именно здесь, в вещественно-волновых измерениях, заложена стратегия волнового программирования развивития и эволюционирования биосистем [1, 2].
Итак, аналогом проявления (образования) голограмм в организме является специфическая динамика хромосомных и других знаковых клеточно-тканевых структур, которые можно рассматривать как амплитудно-фазовые рассеивающие квантованно-динамичные квазиголографические и семантические образования. Одним из законов построения их являются градиенты интенсивности результирующих электромагнитных спекл-полей. Известно, что спекл-поля характеризуют степень модуляции волновых когерентных полей. Для упрощения выкладок ограничимся случаем линейной передаточной функции
![]() (7)
(7)
Подставив выражение (7) в соотношение (6), получим математическую модель амплитудного пропускания клеточно-голографической структуры
![]()
![]() (8)
(8)
![]() Проанализируем
каждый член уравнения (8). Первый член (обозначим его
Проанализируем
каждый член уравнения (8). Первый член (обозначим его![]() )-
фоновый, действительно не содержит никакой информации о фазе волны. Однако, из
кажущегося равномерно-фонового распределения интенсивности поля этого члена еще
не следует отсутствие полезной информации о будущей микро-
и макродисперсной структуре дифференцирующейся живой
клетки или многоклеточного тканевого образования, например, крыла, пальца,
печени, глаза и т.д. Этот фон может выступать как неявный план цитодифференцировок и размещения клеток в
пространстве-времени организмов. Созданный в процессе эмбриогенеза как
промежуточный этап морфо-биохимический фрагмент организма обладает тонкой
амплитудно-фазовой структурой для сканирующих ее эндогенных (созданных в
биосистеме) электромагнитных квантов и акустических полей различных диапазонов.
Такой фрагмент будет выполнять роль динамической
объемно-пространственной квазиспеклголограммы,
“читаемой” электромагнитными и/или акустическими полями с референтными
функциями. Таким путем создаются реестры разметочных волновых фронтов, которые
можно рассматривать как “планы” послойной пространственной и временной
организации биосистемы. Основным биорегуляторным
компонентом этих фронтов (распределенных интенсивностях полей) являются их
градиенты, которые, в свою очередь, могут определять градиенты биоактивных веществ-морфогенов, считавшихся до недавнего времени
главными движущими силами биоморфогенезов в духе
теории диссипативных структур [1]. Но не только морфогенов.
Градиенты могут выступать как прямые силовые факторы, задающие вектора движения
внутриклеточных, клеточных и тканевых структур организма. Вновь созданный
фрагмент организма снова задает пул разметочных фронтов, приводящих к
построению очередной части биосистемы, и так из цикла в цикл.
)-
фоновый, действительно не содержит никакой информации о фазе волны. Однако, из
кажущегося равномерно-фонового распределения интенсивности поля этого члена еще
не следует отсутствие полезной информации о будущей микро-
и макродисперсной структуре дифференцирующейся живой
клетки или многоклеточного тканевого образования, например, крыла, пальца,
печени, глаза и т.д. Этот фон может выступать как неявный план цитодифференцировок и размещения клеток в
пространстве-времени организмов. Созданный в процессе эмбриогенеза как
промежуточный этап морфо-биохимический фрагмент организма обладает тонкой
амплитудно-фазовой структурой для сканирующих ее эндогенных (созданных в
биосистеме) электромагнитных квантов и акустических полей различных диапазонов.
Такой фрагмент будет выполнять роль динамической
объемно-пространственной квазиспеклголограммы,
“читаемой” электромагнитными и/или акустическими полями с референтными
функциями. Таким путем создаются реестры разметочных волновых фронтов, которые
можно рассматривать как “планы” послойной пространственной и временной
организации биосистемы. Основным биорегуляторным
компонентом этих фронтов (распределенных интенсивностях полей) являются их
градиенты, которые, в свою очередь, могут определять градиенты биоактивных веществ-морфогенов, считавшихся до недавнего времени
главными движущими силами биоморфогенезов в духе
теории диссипативных структур [1]. Но не только морфогенов.
Градиенты могут выступать как прямые силовые факторы, задающие вектора движения
внутриклеточных, клеточных и тканевых структур организма. Вновь созданный
фрагмент организма снова задает пул разметочных фронтов, приводящих к
построению очередной части биосистемы, и так из цикла в цикл.
Второй член уравнения (8)
описывает автокорреляцию послойно меняющегося амплитудно-фазового фрагмента
“портрета” организма. Обозначим его через ![]() Третий и четвертый члены содержат
информацию о фазовом распределении потенциальной биоструктуры
в модуляциях фронта образующей волны. Это выражается в комплексном амплитудном
пропускании волн этой же биоструктурой-голограммой.
Соответственно, третий основной член, ранее представленной в виде полевого
плана потенциальной (“планируемой”) биоструктуры, обозначим
Третий и четвертый члены содержат
информацию о фазовом распределении потенциальной биоструктуры
в модуляциях фронта образующей волны. Это выражается в комплексном амплитудном
пропускании волн этой же биоструктурой-голограммой.
Соответственно, третий основной член, ранее представленной в виде полевого
плана потенциальной (“планируемой”) биоструктуры, обозначим![]() а
четвертый (комплексно сопряженный) - через
а
четвертый (комплексно сопряженный) - через![]() .
.
Тогда, при таком обозначении,
амплитудное пропускание квазиголограммы можно
выразить в виде![]() (9)
(9)
Проходя через такую систему квазиголографического фильтра-транспаранта, когерентная волна будет “считывать” имеющуюся на нем информацию как с обычной голограммы. Однако в отличие от обычного считывания, известного в технике голографии, считывание в живом и динамичном организме, вероятно, имеет квантованный по времени характер с возвратами (за счет возвратной памяти в спектрах ФПУ, см. выше), также как квантованными являются движения молекулярных и надмолекулярных структур организма. Последнее отчетливо прослеживается в нелинейной динамике ДНК, хромосом, рибосом и коллагена in vitro - in vivo, по крайней мере, в рамках явления ФПУ [1, 2]. На уровне органов биосистемы квантованность - это прерывистые движение глазного яблока. Квантованность движения субклеточных органелл проявляется в так называемом ядерном циклозе, при котором клеточные ядра нейронов или кариоплазма вращаются. У нейронов вращение ядра происходит по часовой стрелке, затем оно на некоторое время прерывается и продолжается уже в противоположном направлении [8].
В таком случае, даже при
непрерывном прохождении восстанавливающей референтной
волны всегда возможно квантованное по времени когерентное считывание и
когерентный квантованный прием обогащенного информацией волнового фронта,
прошедшего через такую биоквазиголограмму.
В моменты кратковременных остановок вращения при прохождении когерентной волной![]() с квазисферическим волновым фронтом, когда на бесконечно малом
интервале кривая может приближенно аппроксимироваться
прямой, биоголограмма восстанавливается волновым
фронтом, подобным волновому фронту референтной волны,
используемому при голографической записи
с квазисферическим волновым фронтом, когда на бесконечно малом
интервале кривая может приближенно аппроксимироваться
прямой, биоголограмма восстанавливается волновым
фронтом, подобным волновому фронту референтной волны,
используемому при голографической записи
![]() (10)
(10)
Также как и на этапе регистрации ограничимся параболическим приближением:
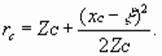 (11)
(11)
Восстанавливающая когерентная
волна (10), проходя через новообразованный слой квазибиоголограммы,
дифрагирует на ее тонкой амплитудно-фазовой
структуре. Образующуюся после этого информационную волну можно описать выражением,
подобным выражению (1), с той лишь разницей, что в фазовом члене будет
присутствовать расстояние![]() . Комплексная
амплитуда дифрагированной биоволны
будет равна
. Комплексная
амплитуда дифрагированной биоволны
будет равна![]() (12)
(12)
причем, ![]() (13) где
(13) где![]() -
расстояние от биоголограммы до плоскости, в которой
мы определяем амплитуду дифрагированной биоволны. Положение точек в этой плоскости определяется
координатой
-
расстояние от биоголограммы до плоскости, в которой
мы определяем амплитуду дифрагированной биоволны. Положение точек в этой плоскости определяется
координатой![]() . Подставив
амплитудный коэффициент пропускания (9) в соотношение (12), получим четыре
члена, из которых первые два определяют закон построения тонкой биоструктуры, а реконструированному изображению, как
волновой схеме потенциальной биоструктуры,
соответствуют два последних члена. Последние проанализируем отдельно. Вначале
рассмотрим третий член, которому соответствует амплитудное пропускание.
. Подставив
амплитудный коэффициент пропускания (9) в соотношение (12), получим четыре
члена, из которых первые два определяют закон построения тонкой биоструктуры, а реконструированному изображению, как
волновой схеме потенциальной биоструктуры,
соответствуют два последних члена. Последние проанализируем отдельно. Вначале
рассмотрим третий член, которому соответствует амплитудное пропускание.
![]() (14)
Подставляя это выражение в соотношение для амплитуды дифрагированной
волны, получим:
(14)
Подставляя это выражение в соотношение для амплитуды дифрагированной
волны, получим:
![]()
Подставив в данное уравнение для![]() в параболическом приближении (2), (4), (11) и (13), после
группировки соответствующих членов, получим
в параболическом приближении (2), (4), (11) и (13), после
группировки соответствующих членов, получим
![]()

![]()
![]()
(16) Это выражение описывает
амплитуду дифрагированной волны, появление которой
обусловлено той частью структуры биоголограммы,
которая приводит к реконструкции основного (или истинного) волнового
изображения того, каким должна быть “планируемая” биоструктура.
Для получения ее точного пространственного прообраза с максимально резким
изображением, биосистеме необходимо начинать строительство потенциальной
структуры на расстоянии, при котором исчезнет квадратичный фазовый член в
уравнении (16), что произойдет при соблюдении следующего условия
![]()
Отсюда следует, что расстояние
основного изображения от биоголограммы определится по
соотношению
![]() (17)
(17)
На расстоянии ![]() получается резкое (высококонтрастное и точное)
изображение потенциальной биоструктуры. Интеграл по
переменной
получается резкое (высококонтрастное и точное)
изображение потенциальной биоструктуры. Интеграл по
переменной ![]()
![]() представляет
одну из возможных форм записи так называемой функции Дирака
представляет
одну из возможных форм записи так называемой функции Дирака ![]() ,
являющейся импульсной функцией, величина которой везде равна нулю, кроме точки
0, где она обращается в бесконечность. Эта функция обладает свойством
,
являющейся импульсной функцией, величина которой везде равна нулю, кроме точки
0, где она обращается в бесконечность. Эта функция обладает свойством
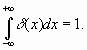 Обозначая в уравнении (16) выражение перед
интегралом через , получим (18):
Обозначая в уравнении (16) выражение перед
интегралом через , получим (18):

![]() так как
так как
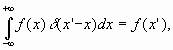 следовательно,
следовательно, ![]() (19)
(19)
где через ![]() обозначено
произведение всех остальных членов. Для сохранения плоскости биоструктуры реконструирующий источник должен находиться на
том же расстоянии от квазибиоголограммы, что и референтный источник, т.е.
обозначено
произведение всех остальных членов. Для сохранения плоскости биоструктуры реконструирующий источник должен находиться на
том же расстоянии от квазибиоголограммы, что и референтный источник, т.е. ![]() При
При ![]() изображение будет находиться на продольной
оси
изображение будет находиться на продольной
оси ![]()
Правильное местоположение восстанавленной биоволны зависит
от взаимного расположения референтных волн по
отношению к биоструктуре, на которой
происходит дифракция.
Теоретический анализ биофизики
управляющих генетико-волновых процессов, проводимый в настоящей работе, а также
ранее [1, 2], приводит к предположению о том, что многоклеточные организмы
обладают высокой избыточностью и помехозащищенностью волновой генетической информации по типу голограмм. Вероятно, хромосомно-водный континуум биосистемы является когерентным
голографическим фракталом, где волновая информация многократно (избыточно)
повторена в разных пространственных масштабах. Отсюда становится понятным,
почему основная часть генома биосистем (гетерохроматин)
устойчива к повреждающим воздействиям. Она голографична.
Другая, малая часть генома (эухроматин), работает,
вероятно, на других информационных принципах, а именно в рамках
семантико-лингвистических построений последовательностей нуклеотидов ДНК и РНК
[2]. И поэтому повреждения эухроматина, как правило,
бывают летальны. Это и понятно. Например, замена только одной буквы в
генетическом “тексте” может полностью поменять смысл “слова”, как это имеет
место при повреждениях генов ферментов. То же относится и к расстановке “знаков
препинания” в ДНК-“текстах”. Однако и здесь роль внутренних и внешних для
биосистемы физических полей значительна. Одной из областей их работы являются
смысловые ориентации выбора омонимов кодонов-антикодонов информационных и
транспортных РНК, которые в свою очередь, определяются контекстными
перестройками хромосомно-волнового континуума
биосистем, а они, перестройки, в какой-то мере, зависят от волнового окружения
земли, выступающего как мегаконтекст (фон) [2]. И в
этом видится объединяющий фрактальный принцип в знаковой системе геном-поле.
Если вернуться к нашей идее, что
хромосомы высших биосистем могут работать также и на принципах квантовой телепортации знаковых фотонных образов, то можно сказать
следующее. Геном нелокален трижды. Один раз по
вещественному измерению, второй и третий - по
волновому. Первое измерение нелокальности имеет
вещественный характер. Это нелокальность голограмм и квазиголограмм хромосомного континуума. В этом же и
фрактальность генома, поскольку любая голограмма - фрактал, т.е. информация в
ней (записанный образ) самотождественна в любом ее
фрагменте, не меньшем, чем длины волн записи. Второе измерение (волновое) - нелокальность волновых фронтов, считанных с таких
голограмм. Каждая область фронта также фрактально повторяет образ, считанный с
голографической решетки. И наконец, третье измерение
(тоже волновое) - это квантовая нелокальность генома,
позволяющая находиться в мгновенном электромагнитном информационном контакте
миллиардам клеточных ядер, благодаря чему организм является суперкогерентной
системой.
Нам предсталяется,
что прием и передача биоголографической (и
радиоволновой) информации может происходить в квантованном по времени
когерентно-импульсном режиме посредством квазизатворов.
Если говорить о фотонных излучениях хромосом, то в качестве квазизатворов
(или шире - переключателей с одной голограммы на другую) могут выступать холестерические послойные жидкокристаллические голограммы
хромосомного континуума. Его холестерики посредством
переориентации директоров меняют направление поляризации проходящего через них
света. Такие регуляторные директора, расположенные ортогонально друг к другу и
ортогонально к распространяющемуся свету, могут пропускать или не пропускать
его за счет поворотов плоскостей поляризации, определяемых направлениями
директоров.
Когерентно-импульсный режим
работы хромосомного континуума как лазерного излучателя может осуществляться по
двум возможным путям. Первый основан на поворотах плоскостей поляризации света
директорами хромосомных холестериков ДНК, а второй
связан с известным явлением по захвату и накоплению фотонов в
слоисто-фрактальных структурах, в том числе вероятно, и в жидких кристаллах
ДНК. При такой ситуации может происходить лазерная накачка фотонов молекулами
ДНК с их последующим квантово-пороговым импульсным отстрелом. Видимо, именно
этот эффект мы наблюдали ранее, фиксируя необычные трапециевидные временные
автокорреляционные функции при динамической лазерной спектроскопии светорассеяния
ДНК и хромосом [1].
В качестве определенного
подтверждения этих положений нами были проведены предварительные
экспериментальные исследования по изготовлению и функционированию специальных ДНК-слоистых устройств типа
“сэндвич” сложного состава, включающего кроме ДНК определенные биометаболиты. Такие устройства при взаимодействии с особым
образом поляризованным лазерным светом со специальной
юстировкой к вектору распространения луча лазера, оказались способными
конвертировать фотоны в биоактивные радиоволны. Такие радиоволны меняют
структуру воды, вводя в нее некую часть информации с ДНК. После этого вода
оказывает четко выраженное действие на рост и развитие растений. Другой
иллюстрацией биоактивности ДНК-промодулированных
радиоволн является эффект оживления невсхожих семян растения Arabidipsis taliana, взятых из чернобыльской зоны и получивших высокую
дозу гамма-облучения. Последний эксперимент служит
некоторым свидетельством в пользу нашей гипотезы о квантовой нелокальности геномов высших биосистем, поскольку какая-то
часть информации с ДНК передавалась параметрически
связанной конверсией фотоны-радиоволны на поврежденный
геном Arabidipsis taliana и регенерировала его. Это наблюдение также
подтверждает полученные нами ранее данные о восстановлении хромосомного
аппарата радиационно поврежденных семян пшеницы и
ячменя с помощью радиоэлектронных устройств, моделирующих радиоволновые
процессы в хромосомах [1].
Таким образом,
мы предложили более развитую био-физико-математическая
модель волнового управления ростом и развитием многоклеточных организмов,
дополняющая и расширяющая первичные, выдвинутые нами ранее [1, 2],
представления в этой области.
ЛИТЕРАТУРА
1.Гаряев П.П. Волновой геном. М.: Изд. Общ. Польза. 1994.
2. Гаряев П.П. Волновой генетический код М.: Издатцентр. 1997.
3.Прангишвили И.В., Ануашвили А.Н., Маклаков В.В.
Закономерности проявления подвижного объекта / Сб. трудов ИПУ РАН. М. Вып. 1. С.7-10.
(1993).
4. Livolant F. Condensed forms of DNA in vivo and in vitro // Physica A. 1991.V.176. P.117-137.
5. Милер М. Голография. Л.: Машиностроение (1979).
6. Гаряев П.П., Тертышный Г.Г., Готовский Ю.В. Трансформация света в радиоволны. III международная конференция “Теоретические и клинические аспекты применения адаптивной резонансной и мультирезонансной терапии”. “ИМЕДИС”. Москва. 18-20 апреля 1997г. с.303-313. (1997).
7. Гаряев П.П., Тертышный Г.Г., Лощилов В.И., Щеглов В.А., Готовский Ю.В. Явление перехода света в радиоволны применительно к биосистемам. Сборник научных трудов “Актуальные проблемы создания биотехнических систем”. Выпуск 2. МГТУ им. Н.Э.Баумана. Академия Медико-Технических Наук РФ. Москва. С.31-42. (1997).
8. Движение немышечных
клеток и их компонентов. Академия Наук СССР. Л.: Наука, 1977. С.105, 167.
9. Einstein A., Podolsky
B., Rosen N., Can quantum-mechanical description of physical reality be considered complete? // Phys.Rev.
1935. V.47. P.777-780; Bouwmeester D., Pan Jian-Wei, Mattle K., Eibl M., Weinfurter H., Zeilinger A. Experimental quantum teleportation // Nature. 1997.
V.390. P.575-579.
Петр Гаряев, д.б.н., акад. РАМТН и РАЕН
Проведенные исследования, казалось бы, не связаны. Однако это не так. Процесс образования фантомов ДНК был обнаружен нами еще в 1984 г. Обнаружение и регистрация волновых реплик ДНК in vitro подтвердила эти результаты. Это еще раз подтверждает нашу гипотезу о способности ДНК быть структурой волнового автореплицирования и автосканирования внутриклеточного метаболического статуса, а в итоге всего организма с целью биокомпьютерного регулирования жизнедеятельности. Это полностью соответствует основному положению нашей концепции «волнового генома»: хромосомный континуум работает как стратегическая знаковая система на 2-х уровнях — вещества и поля одновременно, реализуя свои функции в качестве квазиразумной системы.
Однако это не единственная волновая система саморегуляции многоклеточных организмов. Вторая система основывается на когерентных излучениях хромосом в Уф-, видимой и ИК- областях спектра. Это дает возможность хромосомному аппарату использовать принципы голографической свертки-развертки генетической информации, квантовую нелокальность ее, а также лингвистические особенности работы дуэта «рибосома-иРНК» в рамках понятия квазиразумности генома как биокомпьютера..
Сейчас сложилась парадоксальная и многообещающая ситуация в генетике, молекулярной биологии и, соответственно, в медицине. Изучен геном человека. На это понадобились 10 лет усилий программы «Геном человека», и теперь известна последовательность всех букв в нашем генетическом тексте.
Трансгенная инженерия набирает силу, и уже множество растений, животных и бактерий являются носителями искусственно внедренных в них полезных для человека генов. Сделаны первые шаги в клонировании животных с перспективой получить первый клон человека.
Но вот парадокс — чем выше наши технологические успехи в генетике и молекулярной биологии, тем дальше мы от всеобъемлющего понимания принципов работы генетического аппарата. Все несомненные успехи в этой области преимущественно связаны с функциями генов, которые кодируют белки. Эти гены занимают около 2% всей генетической памяти хромосом. Остальная, главная часть, т.е. 98%, до сих пор является непонятой генетиками и, вероятно, поэтому они назвали ее «мусором». Есть гипотезы, но все они призваны оправдать существование «мусорной» ДНК как некоего помощника для 2% «кодирующей» ДНК. Либо «мусор» трактуют как «кладбище вирусов»(!)
Игнорировать или так наивно понимать роль 98% генома — явная ошибка. Более того, правильно ли мы видим истинную роль 2% генетической информации, если 98% её для нас фактически — «terra incognita»? Понимаем слабо, поскольку не можем полностью излечивать рак, не можем противостоять ВИЧ, не можем бороться против туберкулеза, не можем продлить жизнь людей, по крайней мере, до рубежа 200 лет и т.д. Обещания генетиков обернулись опасными для жизни трансгенными продуктами питания, грозящим дисбалансом биосферы.
Клонирование животных дает нам, в основном, тщательно скрываемых уродов, или животных, которые аномально быстро стареют и умирают, как знаменитая овечка Долли.
Вполне естественно, что научное сообщество, например, в лице большой группы шведских ученых, начинает бить тревогу по этому поводу http://www.psrast.org/defknthe.htm Где выход из этого странного состояния обилия экспериментального и противоречивого материала и дефицита достаточно полного теоретического осмысления принципов работы хромосом? Та же группа шведских ученых полагает, что одним из прорывных научных направлений являются наши разработки http://www.rialian.com/rnboyd/dna-wave.doc
Суть наших идей, и что они дают практике, сводится к следующему. Мы исходим из очень простых стратегических соображений. Для успеха в попытках кардинально лечить людей и резко затормозить процессы их старения надо понять языки, на которых клетки общаются друг с другом. В какой-то степени нам удалось это сделать. Оказалось, что это языки тех самых 98% «мусора» в нашем собственном генетическом аппарате http://www.self-managing.net/genetica/Zip/V_Genetica%20(Kniga).zip
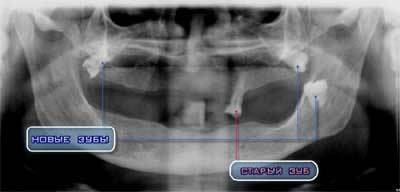
Уникальный рентгеновский снимок ротовой полости
пациентки 60 лет – зубы, выросшие с наклоном
под постоянно носимым протезом
Основным из них является язык голографических образов http://www.self-managing.net/genetica/Zip/Nestazionarn_golografia.zip , основанный на принципах лазерных излучений генетического аппарата http://www.self-managing.net/genetica/Zip/DNK%20laser.zip , работающего как квазиразумная структура http://www.rialian.com/rnboyd/dna-wave.doc .Важно, что нашему генетическому аппарату свойственны реальные процессы, существенно дополняющие триплетную модель генетического кода http://www.self-managing.net/genetica/Zip/Central%20Dogma.zip
. В этих процессах используются ранее неизвестные для организмов свойства http://www.self-managing.net/genetica/Zip/FROL.zip и др.Что нам дают эти новые знания? Ответ таков: теперь, зная об этом и имея физико-математическое описание дополнительных информационных процессов в генетическом аппарате, мы создали аппаратуру, которая моделирует волновые информационные функции живой клетки и ее генетического аппарата. Устройства такого рода являются фактически первыми квантовыми биокомпьютерами. Они позволили осуществить дальнюю многокилометровую передачу генетико-метаболической информации в форме особых электромагнитных полей, введение ее в биосистему-акцептор и стратегическое управление в отношении ее, биосистемы, биохимического и физиологического состояний http://www.self-managing.net/genetica/Zip/SFR.zip . В частности, удалось регенерировать эндокринные железы у животных, а также резко затормозить старение людей.
Эл.адрес: gariaev@mail.ru
Интернет-сайт: http://www.self-managing.net/genetica
Волновые репликативные отображения ДНК и ее ближайшего окружения
П.П.Гаряев, Валтрауд
Вагнер, Е.А.Леонова-Гаряева, А.Е.Акимов,
И.А.Сухоплюева, Э.Р.Мулдашев
В этой работе мы показываем, что воздействие на препарат ДНК in vitro определенными
сочетаниями электромагнитных полей в диапазоне УФ-ИК вызывает ответ в форме
многообразных, динамичных, долгоживущих реплицирующихся волновых отображений
ДНК и ее ближайшего окружения.
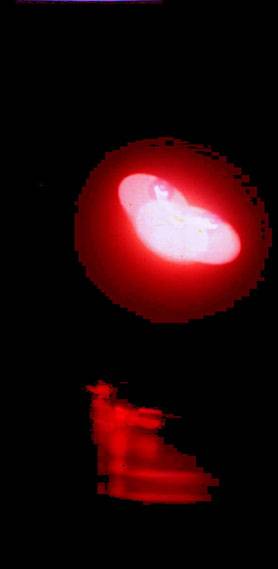
Мы обнаружили способность препаратов ДНК in vitro давать многократно реплицирующиеся отклики-отображения, реплики самой ДНК и ее ближайшего окружения. Такие реплики являются откликом на определенные комбинации воздействия электромагнитных полей в диапазоне УФ-ИК. Реплики могут выстраиваться строго по горизонтали в обе стороны, двигаться по сложным траекториям, а также сохраняться какое-то время после прекращения воздействия полей на препарат ДНК. (Рис.1-4).

Рис.1. Эффект волнового реплицирования ДНК и ближних окружающих предметов. Слева
контроль, справа опыт. Второй вариант методов выявления волновых реплик ДНК.


Рис.2 (a). Дискретность и сложность траектории волновых реплик ДНК. (слева – исходное изображение, справа – контрастированное). Первый вариант методов выявления волновых реплик ДНК.



Рис.2 (б). Распределение яркостей изображения по каналам цветности: красный (R), зеленый (G), синий (B).


Рис.3 (a). Пространственная динамичность волновых реплик ДНК. Видна тонкая продольная структура траектории трека.



Рис 3 (б).Распределение яркостей по каналам цветности: красный (R), зеленый (G), синий (B).
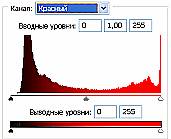
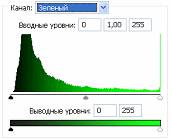
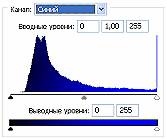
Рис 3.2. Гистограммы значений яркостей по каналам цветности: красный, зеленый, синий.
На контрольных снимках, когда источники воздействующих на биопрепарат физических полей не включены, данного явления не наблюдается. Также как нет эффектов при действующей аппаратуре, но без ДНК. За время экспозиции (1 сек.) в первом варианте экспериментов (см. Методы) волновые реплики ДНК перемещаются в пространстве и мультиплицируются, их траектория не предсказуема и дискретна. Генерация реплик происходит только при условии взаимной пространственной стационарности препарата ДНК, источников возбуждающих полей и фотоаппарата. Прикосновение к препарату ДНК (второй вариант экспериментов) вызывает изменение вектора распространения реплик в пространстве на диаметрально противоположное (слева направо), а затем исчезновение (Рис.5 (а, б, в)). Существенно, что в ряде случаев имеет место не только волновое мультиреплицирование препарата ДНК, но и окружающих предметов (Рис. 1, 5 (а, б)).
Для анализа цветного отпечатка на эмульсии фотопленки приведены изображения, полученные при сканировании пленки, как распределение яркостей в разных каналах цветности: красном (R), зеленом (G), синем (B): на Рис. 2(б), 3(б), 4(б). В изображении, представленном на Рис. 2 (а, б), видно, что почти вся яркостная шкала изображения реплик находится в красном канале цветности, в зеленом и синем они слабо различимы. Такое же превалирование яркостного диапазона красного канала хорошо видно на сопровождающих Рис.3 (б) гистограммах яркостей (Рис. 3 (в)) соответствующих каналов. Обращает на себя внимание именно та часть гистограммы, где расположено реплицированное изображение: участок яркостей в диапазоне значений от 128 до 255 – хорошо отличаемый пик значений в этой части гистограммы красного по сравнению с почти равномерным убыванием в такой же области гистограмм зеленого и синего каналов. Возможно, такое распределение яркостей может указывать на то, что сфотографированный дискретный трек есть опосредованная препаратом ДНК многократно повторенная реплика световой матрицы аппарата «Дюна-М» красного и инфракрасного цветов (позиция 8 на Рис. 8), на которой находится препарат ДНК в момент генерации реплик. Этот источник света представляет матрицу из 37 чередующихся диодов, из них красных – 21, инфракрасных – 16. В эксперименте препарат ДНК был помещен в пробирку эппендорф и перекрывал 5 диодов. Это коррелирует с 5-кратной продольной исчерченностью трека реплики (Рис. 3a).
Важным свойством волновых реплик ДНК является относительная долговременность их существования после выключения всех инициирующих физических полей (Рис. 4), но время жизни реплик может зависеть также от чувствительности фотопленки, фиксирующей реплики. А также от ее спектральной избирательности.

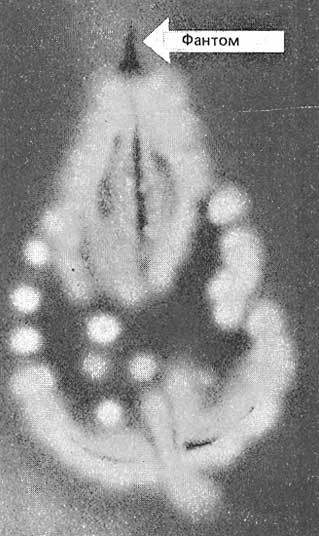
Рис. 4 (a). Волновая долгоживущая реплика ДНК
из эксперимента на Рис. 3 («фантом») после выключения возбуждающих электромагнитных полей.



Рис. 4 (б). Распределение яркостей по каналам цветности: красный, зеленый, синий.
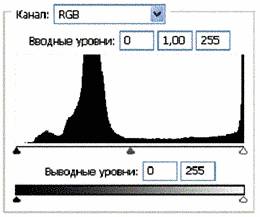
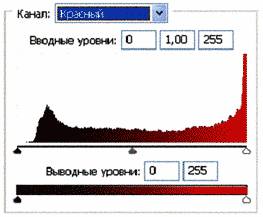
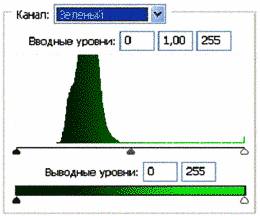
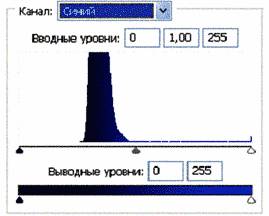
Рис. 4 (в).Гистограммы. Распределение яркостей по каналам цветности: красный, зеленый, синий.
Первая слева в верхнем ряду: гистограмма RGB – общая (совмещенная).
Можно заметить существенное отличие в распределениях яркостей по каналам цветности у изображения волновой реплики ДНК и «фантома» этой реплики, остающегося после выключения источников света. Анализ изображений на Рис. 4 (б) и сопоставление его с гистограммой на Рис.4 (в) выявляет то, что изображение «фантома» реплики в красном канале становится размытым как по диапазону, так и по амплитудам значений яркостей. При этом в зеленом и синем каналах наблюдается сужение локализации значений в диапазоне от 70 до 120, виден четкий пик, что объясняет яркость реплики ДНК, ее оттенки и полутона, особенно, в зеленом канале.

Рис.5 (а). Момент механического воздействия на препарат ДНК. Второй вариант методов выявления волновых реплик ДНК.

Рис. 5 (б). Сдвиг волновых реплик налево сразу после механического воздействия. Характерно появившееся резкое деление кадра по свето- и цветопередаче, не связанные со стабильной работой шторки фотоаппарата.

Рис. 5 (в). Исчезновение эффекта образования реплик ДНК через 5-8 сек. после механического воздействия на препарат ДНК. При этом вся аппаратура, инициирующая реплики, остается включенной.
Тип образующихся реплик в каждом конкретном случае в рамках используемых методов стохастический (1-й вариант методов) или детерминированный (2-й вариант) в пространстве-времени.
Одним из результатов по индуцированию волновых реплик окружающих предметов с помощью препарата ДНК по первому методу явилось мультиплицирование тройного изображения лампы БС (УФ-В) (Рис. 6).
|
|
|
|
Рис.6(а). Мультиплицирование тройного изображения лампы БС. Оригинал. |
Рис.6(b).
Мультиплицирование тройного изображения лампы БС. Контрастированное
изображение |
В качестве одного из контролей на способность генерировать волновые реплики использовали хлорид натрия (кристаллический); хлорид натрия (1М водный раствор); винная кислота в кристаллах; винная кислота рацемат (1М водный раствор); крахмал воздушно сухой; глицин кристаллический; кальциферол воздушно сухой; токоферол воздушно сухой; хлорофилл воздушно сухой; вода бидистиллированная; интерферон в смеси с Bacillus subtilis, воздушно сухие. Ни один из препаратов не дал реплик.
|
|
Рис. 7. Матрица с красными и инфракрасными диодами (аппарат «Дюна М» или «Дюна Т». http://argonet.ru/nar_lechebn_duna.htm ) Содержит 37 диодов,
из них красных – 21 ( |
Методы
Мы использовали две схемы экспериментов. Первая приведена на (Рис.9), вторая достаточно проста и видна напрямую на фото (Рис. 5(в)). Схемы также даны в18. Для получения и визуализации волновых репликативных отображений ДНК по первой схеме производят следующие операции. Посредством реле времени (позиция 3 на Рис.8) в разных сочетаниях включаются необходимые излучатели БС (УФ-В, представляющий лампу накаливания синего цвета «ЛОН БС 75», из стекла СЛ98-1) (позиция 5) и матрица с красными и инфракрасными диодами (Рис. 8)) (позиция 8 на схеме Рис. 8), а также лампа ртутная бактерицидная, или лампа Compact electronic CEST26E27 Black (УФ-С) (позиция 6), или БС (УФ-В) и МХТ-90 (холодный катод (позиция 4)). При включении источников света воздушно сухой препарат ДНК из селезенки быка (РЕАХИМ, марка А, НПО «Биолар») (около 100 мг в закрытой пластиковой конической пробирке «эппендорф» длиной 4см, диаметром в верхней части 0,9см) или (3мл водного раствора ДНК, 1мг/мл) находится в зоне действия излучателей (1мм-50см от источников света). Ход работы снимается на фотоплёнку Fuji 24-27 DIN. При этом работает осциллограф (позиция 2) с электродами, которые регистрируют электромагнитные поля в зоне проведения эксперимента и фиксируется усреднённый нормальный электромагнитный фон в помещении, определяемый по характеру синусоиды на осциллографе. Далее посредством реле времени через 10 мин. отключают излучатель УФ-С. Фотографически обнаруживается возникновение динамичных специфических волновых структур, невидимых глазами, но фиксируемых на фотопленке мультиплицированных реплик ДНК и ближних окружающих объектов, непосредственно связанных с фотонным воздействием на ДНК. Иначе говоря, происходит умножение количества отображений препарата ДНК, распределяемых в пространстве по сложным траекториям (1-й вариант методов) и по горизонтали (2-й вариант методов), а также отображений объектов, связанных с возбуждением ДНК.
Второй вариант методов получения и визуализации волновых репликативных отображений ДНК (Рис. 5(в)) состоит в следующем. Воздушно сухой препарат ДНК, 100мг, помещают в открытом виде в держатель из алюминиевой фольги. С интервалом 2-3 сек. включают лампу БС (УФ-В), лампу сompact electronic CEST26E27 Black (УФ-С) и аппарат «Дюна М». Через 5 мин. начинают фотосъемку на фотоплёнку Fuji 24-27 DIN. В этом варианте регистрируются реплики ДНК и ближних объектов, которые распространяются строго направо. При механическом воздействии на препарат ДНК вектор распространения реплик меняет своё направление на диаметрально противоположное, то есть налево. Затем через 5-8 сек. после механического воздействия, несмотря на то, что вся аппаратура, инициирующая реплики, остается включенной, реплики исчезают (или не фиксируются используемым типом фотопленки).

Обсуждение
Несмотря на успехи молекулярной генетики и клеточной биологии, до сих пор тонкие механизмы знаковых функций генетического аппарата остаются непонятными. Особенно ярко это проявилось после публикации работы, в которой команде Pruitt удалось изящно доказать, что законы Менделя в генетике не всегда выполняются, более того, поведение одного из генов не подчиняется здравому смыслу1. Это проявилось в том, что во взрослых растениях Arabidopsis фенотипически проявился нормальный ген Hothead, которого не было в исходных мутантных по этому гену семенах. Нормальный ген Hothead был у предков растения. Отсутствующий в хромосомах посеянных семян ген Hothead в 10% случаев заменил реальный мутантный ген hothead, когда эти семена дали взрослые растения. Объяснения этому феномену пока не дано. Высказано предположение, что нормальный ген хранился как его ревертазная РНК копия. Это слабое и уязвимое объяснение, не имеющее экспериментального доказательства. Необъяснимый с позиций классической генетики феномен возврата предкового гена поднял на поверхность целый ряд нерешенных кардинальных вопросов генетики и эмбриологии. Их можно кратко перечислить:
a/ воблирование 3-го нуклеотида в кодоне делает каноническую
таблицу генетического кода ареной потенциальных ошибок в синтезе белков поскольку
она автоматически даёт омонимии значащих дублетов в кодонах, когда пары
одинаковых дублетов кодируют разные аминокислоты2; при
этом третьим нуклеотидом кодона может быть любой из четырех, что постулировал
F.Крик3; тем не менее, ошибки при выборе аминокислот чрезвычайно маловероятны,
и это удивительный факт; b/ ситуация, когда 98%
генома эукариот считают «мусором», в лучшем случае, помощником триплетного
кода, или «кладбищем» вирусных геномов; c/ 2%
кодирующей ДНК человека (около 40тыс. генов) оказалась весьма близкими таковым
свиней, ослов, мух и даже E.coli; d/
гены по непонятным причинам и законам транспозицируются
в 3-D континууме хромосом; e/ гены по непонятным
причинам разбиты на интроны и экзоны;
f/ матрично независимый синтез РНК последовательностей Q ![]() репликазой
бактериофага E. coli и такой же синтез РНК
последовательностей РНК полимеразой бактериофага T7 E. coli4,5, которые
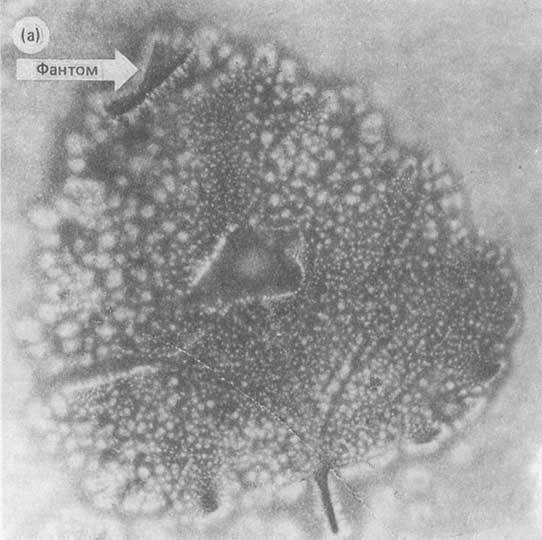
нарушает канонический принцип чисто вещественной репликации ДНКРНК; g/ фантомный листовой эффект, когда часть живого листа
растения воспроизводит его целый образ в режиме газо-разрядной (Кирлиановской) визуализации6,7; h/ непонятные по механизмам дистантные
функции селекторных генов морфогенеза, когда их продукты синтезируются в одном
месте, а действие сразу проявляется в другом, далеко отстоящем месте
развивающегося эмбриона.
репликазой
бактериофага E. coli и такой же синтез РНК
последовательностей РНК полимеразой бактериофага T7 E. coli4,5, которые
нарушает канонический принцип чисто вещественной репликации ДНКРНК; g/ фантомный листовой эффект, когда часть живого листа
растения воспроизводит его целый образ в режиме газо-разрядной (Кирлиановской) визуализации6,7; h/ непонятные по механизмам дистантные
функции селекторных генов морфогенеза, когда их продукты синтезируются в одном
месте, а действие сразу проявляется в другом, далеко отстоящем месте
развивающегося эмбриона.
Все эти и другие мало понимаемые проявления знаковых функций хромосом заставляют думать и доказывать, что генетическая память имеет иные атрибуты, существенно дополняющие белковый код. Вероятно, эти атрибуты имеют волновую природу. Именно эти противоречия и неполнота модели триплетного белкового кода для объяснения кодирования пространственно-временной структуры многоклеточных биосистем заставила нас обратить внимание на работу генетического аппарата как системы высоко организованных знаковых излучений электромагнитных и акустических полей8,9,10,11. Вероятно, одним из проявлений волновых знаковых функций ДНК являются обсуждаемые здесь результаты нашего исследования по волновым репликам ДНК. Первые свидетельства, что ДНК способна к волновому типу памяти были получены нами в 1985 г. Мы обнаружили феномен так называемой ДНК фантомной памяти (DNA phantom effect)12и подробно описали его в работе8. Возможно, он имеет прямое отношение к излагаемым здесь результатам. ДНК фантомный эффект проявляется следующим образом. При работе с ДНК, как линейным полимером, с использованием метода динамического лазерного светорассеяния (ДЛС), в кюветном отделении спектрометра остаются некие гипотетические структуры (объекты), специфическим образом рассеивающие свет после удаления кюветы с препаратом ДНК. Именно это явление было названо ‘DNA phantom effect’. Контрольные замеры, до помещения ДНК в спектрометр, дают только фоновое светорассеяние. Продувание кюветного отделения прибора газообразным азотом приводит к исчезновению фантома, однако он вновь регистрируется спустя 5-7 минут. Фактически близкий эффект наблюдали Allison et. al.13, которые также использовали ДЛС для изучения рестриктных фрагментов ДНК. Они обнаружили аномальное светорассеяние таких фрагментов. Авторы определили этот феномен как ‘mimicking the effect of dust’, то ест как эффект, имитирующий пыль. В их исследовании этот эффект не являлся результатом пылевых загрязнений, но следствием влияния ‘объекта’, который вел себя как пылеподобные ‘частицы’, рассеивающие свет. Поведение таких ‘частиц’ задает такие характеристики светорассеяния, которые не согласуются с классическими моделями процесса светорассеяния для линейных полимеров14,15,16. Между экспериментами8,12,13есть общие моменты, связанные с тем, что на препараты ДНК воздействовали светом в видимой области – 632,8нм и 488нм. То же самое имеет место при инициации волновых реплик ДНК в данной работе. В образовании волновых реплик ДНК доминирующую функцию берёт на себя, вероятно, тот излучатель, длина волны которого совпадает с длиной волны поглощения ДНК. Это источник УФ света.Мы не можем исключить, что в экспериментах8,12,13возникали волновые реплики ДНК, которые искажали классическое светорассеяние для линейных полимеров. Мы не можем также исключить, что волновые реплики ДНК могут играть определенную роль в процессах волновых генетических коммуникаций между клетками в онтогенезе и во взрослом состоянии организмов.
Для волновых реплик ДНК in vitro свойственен квази генетический процесс: ДНК волновым путем копирует самою себя и объекты (устройства, оборудование), вовлеченные в инициацию реплицирования (Рис. 1-3, 5 (a), (b); 6 (a), (b)). Этот момент поведения генетического аппарата in vivo у многоклеточных организмов является одним из ключевых в нашей модели волнового генома8,9,10,11. В соответствии с этой моделью, каждая клетка и вся биосистема в целом осуществляет непрерывное сканирования своего генетико-метаболического состояния. Иными словами, ДНК за счет собственных когерентных излучений in vivo поляризационно-голографическим образом19считывает самою себя и внутриклеточное метаболическое пространство в диапазоне 250-800нм, то есть копирует, создает волновые реплики своего структурно-функционального статуса в каждый момент времени. Возможно, диапазон частот гораздо шире, но технически пока возможна регистрация только указанного спектра частот. Такие реплики образуют так называемое спутанное (entanglement) состояние и нелокально (и мгновенно) информационно объединяют биосистему20.
Вместе с тем, мы должны подчеркнуть, что обнаруженные эффекты образования волновых реплик ДНК нуждаются в более точной экспериментальной постановке. Очень многое в этой части остается неясным и поэтому исследования продолжаются.
ТЕОРЕТИЧЕСКОЕ ПРИЛОЖЕНИЕ
В течение длительного времени физика оперировала четырьмя взаимодействиями, — сильными и слабыми взаимодействиями, электромагнитными и гравитационными полями. С помощью этих взаимодействий удавалось объяснить почти все известные явления природы и многочисленные лабораторные эксперименты. Однако постепенно стала накапливаться экспериментальная феноменология, которую не удавалось объяснить с помощью указанных взаимодействий21. Методологически было понятно, что такое положение свидетельствовало о неполноте наших знаний о природе. Для понимания экспериментальной феноменологии надо было сформулировать новую научную парадигму. Первые шаги в этом направлении были сделаны Э.Картаном, который постулировал существование в природе полей, порождаемых плотностью углового момента вращения22. Эти поля были названы им полями кручения или торсионными полями. Многочисленные исследования в этом направлении в 70-е годы ХХ столетия привели к созданию Теории Эйнштейна – Картана (ТЭК), которая связывала природу кручения с гравитацией. Позже было показано, что торсионные поля порождаются спином, в отличие от электромагнитных полей, порождаемых зарядом, и гравитационных полей, порождаемых массой. В этом случае торсионные поля рассматриваются как самостоятельная физическая реальность, как пятое взаимодействие, «пятая сила».
В 80-е годы ХХ столетия А.Е.Акимовым была построена фитонная модель физического вакуума21, которая в период до создания теории физического вакуума Г.И.Шиповым23позволила понять физические механизмы, связанные с действием торсионных полей. При построении моделей физического вакуума было признано целесообразным вернуться к электронно-позитронной модели П.Дирака в несколько измененной ее интерпретации. Учитывая, что вакуум определяется как состояние без частиц, и, исходя из модели классического спина Я.Френкеля и И.Тернова24,25как кольцевого волнового пакета (следуя терминологии Белинфанте — циркулирующего потока энергии), будем рассматривать вакуум как систему из кольцевых волновых пакетов, соответствующих электронам и позитронам, а не собственно электронно-позитронным парам, несущим информацию о свойствах этих частиц (рис. 1).
При сделанных предположениях нетрудно видеть, что условию истинной электронейтральности такого полевого электронно-позитронного вакуума будет отвечать состояние, когда кольцевые волновые пакеты электрона и позитрона будут вложены друг в друга. Если при этом спины этих вложенных кольцевых пакетов противоположны, то такая система будет самоскомпенсирована не только по зарядам, но и по классическому спину и магнитному моменту. Так как масса покоя волновых пакетов равна нулю, то такие системы будут обладать гравитационной нейтральностью. Такую систему из вложенныхкольцевых волновых пакетов будем называть фитоном. Фитонный континуум образует материальную среду – физический вакуум. Рассмотрим наиболее важные в практическом отношении случаи возмущения разными внешними источниками физического вакуума в рамках построенной модели.

Рис.1. Фитонная структура физического
вакуума и его поляризационные состояния.
1. Пусть источником возмущения является заряд — q. Если вакуум имеет фитонную структуру, то действие заряда будет выражено в зарядовой поляризации физического вакуума. Это состояние фиксируется как электростатическое или электромагнитное поле. Этот случай хорошо известен в квантовой электродинамике. В частности, Лэмбовский сдвиг традиционно объясняется через зарядовую поляризацию электронно-позитронного физического вакуума.
2. Пусть источником возмущения является масса — m. В отличие от предыдущего случая, когда мы столкнулись с общеизвестной ситуацией, здесь будет высказано гипотетическое предположение. Возмущение физического вакуума массой m будет выражаться в симметричных колебаниях элементов фитонов вдоль оси на центр объекта возмущения. Такое состояние физического вакуума может быть охарактеризовано, как гравитационное поле (G-поле).
3. Пусть источником возмущения является классический спин — s. Будем предполагать, что действие классического спина на физический вакуум будет заключаться в следующем. Если источник имеет спин, то спины фитонов, которые совпадают с ориентацией спина источника, сохраняют свою ориентацию. При этом спины фитонов, которые противоположны спину источника, под действием источника испытают инверсию. В результате физически вакуум перейдет в состояние поперечной спиновой поляризации. Это поляризационное состояние можно интерпретировать как спиновое поле (S — поле) — поле порождаемое классическим спином, т.е. торсионное поле, — в другой интерпретации дальнодействующее спинорное поле, если учесть, что торсионное поле может быть представлено через пару спинорных полей. Сформулированный подход созвучен представлениям о полях кручения, как конденсате пар фермионов.
Поляризационные спиновые состояния SR и SL противоречат запрету Паули. Однако согласно концепции М.А.Маркова26, при плотностях порядка планковских, фундаментальные физические законы могут иметь другой, отличный от известных вид.
В соответствии с изложенным подходом можно говорить, что единая среда – физический вакуум может находиться в разных фазовых, точнее, поляризационных состояниях — EGS состояниях. Эта среда в состоянии зарядовой поляризации проявляет себя как электромагнитное поле (Е). Эта же среда в состоянии спиновой продольной поляризации проявляет себя как гравитационное поле (G). Наконец, та же среда — физический вакуум в состоянии спиновой поперечной поляризации проявляет себя как спиновое (торсионное) поле (S). Таким образом, ЕGS — поляризационным состояниям физического вакуума соответствуют EGS -поля.
В начале 80-х годов в работах А.Е.Акимова было предсказано и экспериментально подтверждено существование особого класса торсионных полей — электроторсионных полей21. В середине 90-х годов автор теории физического вакуума Г.И.Шипов дал строгое на фундаментальном уровне обоснование электроторсионных полей22.
Модель поляризационных состояний физического вакуума позволила установить весьма важное обстоятельство. Уже отмечалось, что торсионное поле фиксируется в случаях, когда физический вакуум переходит в состояние спиновой поперечной поляризации. Однако можно сформулировать более общий подход, опираясь на критерии синергетики. Будет считать, что торсионное поле фиксируется всегда, когда физический вакуум находится в спиново неравновесном состоянии.
Нетрудно видеть, что при зарядовой поляризации физического вакуума зарядовое расщепление фитонов приводит и к спиновому пространственному расщеплению. В результате спины оказываются некомпенсированными, что будет равносильно появлению торсионной компоненты в электромагнитном поле. Если гравитационные и торсионные поля проявляются в «чистом виде», то электромагнитные поля всегда содержат торсионную компоненту, что является важным фундаментальным фактом. Торсионное поле наблюдается как в электростатическом поле, так и в электромагнитном излучении.
С изложенных позиций нетрудно предположить, что при электромагнитном или звуковом воздействии на субстрат ДНК создаётся его зарядовая неравновесность. Следствием этого является возникновение электроторсионного поля. В рассмотренных выше экспериментах неотвратимо практически все объекты становились такими источниками. При этом первичным объектом возмущения был субстрат ДНК, который своим электроторсионным полем индуцировал возмущение других объектов эксперимента.
А) 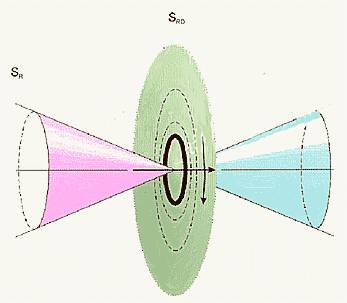
B) 
Рис.2. Диаграмма направленности
торсионного поля спинирующего источника (а)
и зависимость интенсивности
статического торсионного поля от расстояния (б).
Теперь рассмотрим диаграмму
направленности источника торсионного поля, в частном случае, — объект со спином
или с угловым моментом вращения для макроскопических тел (рис. 2). На
расстояниях больших размера источника электроторсионного
поля это поле имеет периодическую структуру. Поэтому на каждом интервале такой
периодической структуры будет отображение источника торсионного поля. В
зависимости от знака поля внешнего воздействия будет отображаться, либо правая,
либо левая часть таких периодических структур. Возможно, именно этот эффект мы
наблюдаем на Рис. 5 (а, б) (экспериментальная часть), где эффект правых реплик
ДНК и ее окружения меняется на левые реплики при механическом воздействии на
препарат ДНК.
При этом остаётся вопрос, как это
всё может фиксироваться на фотографиях, если учесть, что фотоплёнка фиксирует
изображения фотохимическим способом, т.е. за счёт электромагнитных воздействий,
а сами исходные структуры имеют торсионную природу. Возможность фиксации на
плёнках торсионных пространственных структур связана со следующими факторами. В
фотографических устройствах (фотоаппаратах с фотоплёнкой) фотоны, попадая в
эмульсию, вызывают фотохимические реакции. В результате при последующих
химических реакциях проявления и закрепления на фотоплёнке появляется
изображение. При действии торсионных излучений на фотоактивные
вещества эмульсии фотоплёнки изменяется спиновое состояние этих веществ.
Результат изменения спинового состояния атомов приводит к химическим изменениям
как следствие изменения взаимодействия валентных электронов, которое подобно тем
изменениям, которые наблюдаются при действии на эти вещества фотонов. Это и
позволяет получать фотографии пространственных структур, которые обсуждаются в
данной статье.
Физическая интерпретация
наблюдаемых явлений, представленных выше, базировалась на важных, но частных
эффектах проявления торсионных полей. Рассмотрим указанную экспериментальную
феноменологию с позиций основных свойств торсионных полей. Укажем на наиболее
важные свойства торсионных полей.
1. Источником торсионных полей
является классический спин или макроскопическое вращение. Торсионные поля могут
порождаться кручением пространства или порождаться как следствие возмущения
физического вакуума, которое имеет геометрическую или топологическую природу, а
также возникать как неотъемлемая компонента электромагнитного поля. Торсионные
поля могут самогенерироваться.
Во всех указанных случаях речь
идет о торсионных полях, порождаемых на уровне вещества. Однако согласно теории
физического вакуума существуют первичные торсионные поля, которые порождаются
Абсолютным Ничто. Подобно тому, как исходный материал мира вещества —
элементарные частицы — рождаются из физического вакуума, в свою очередь
физический вакуум рождается из первичного торсионного поля.
2. Квантами торсионного поля
являются тордионы. Есть основания считать, что тордионами являются низкоэнергетические нейтрино с энергией
порядка единиц эВ.
3. Так как торсионные поля
порождаются классическим спином, то и при их воздействии на те или иные объекты
у этих объектов в результате воздействия может измениться только их спиновое
состояние (состояние ядерных или атомных спинов, а также спинов валентных
электронов).
4. Торсионные поля обладают
аксиальной симметрией относительно своего источника.
5. Торсионные поля (Т),
порождаемые классическим спином, могут быть аксиальные (Тa) и радиальные (Тr). Каждое из этих торсионных
полей может быть правое (ТaR, ТaL)
или левое (TaL,
ТrL).
В общем случае торсионные поля обладают аксиальной симметрией.
6. Одноименные
торсионные заряды (одноименные классические спины (SRSR или SLSL) притягиваются, а разноименные (SRSL) — отталкиваются.
7. Стационарный спинирующий объект создает статическое торсионное поле.
Если у спинирующего объекта есть какая-либо неравновесность: изменение угловой частоты, наличие у объектов прецессии, нутации или моментов более высокого
порядка, неравномерного распределения масс относительно оси вращения для
массивных объектов, то такой динамический спинирующий
объект создает волновое торсионное излучение.
8. Статическое торсионное поле
имеет конечный радиус действия rо, на интервале которого
интенсивность торсионного поля слабо варьирует. Условно по аналогии с
электромагнетизмом, хотя физика процессов здесь другая, этот интервал rо
можно назвать ближней зоной. Волновое торсионное излучение не ограничено
интервалом rо
и его интенсивность не зависит от расстояния.
9. Средой, через которую
распространяются торсионные излучения, является физический вакуум. По отношению
к торсионным волнам физический вакуум ведет себя как голографическая среда. В
этой среде торсионные волны распространяются через фазовый портрет этой
голограммы. Этот основополагающий физический фактор объясняет информационный
(не энергетический) характер передачи сигналов, а также сверхсветовую скорость
передачи сигналов.
10. Для торсионных полей
потенциал тождественно равен нулю, что соответствует их неэнергетическому
характеру. Это второй фактор, который определяет
почему торсионные сигналы (воздействия) передаются информационно, а не
энергетически, т.е. без переноса энергии.
11.Константа спин-торсионных взаимодействий для статических
торсионных полей с кручением Картана по существующим оценкам меньше, чем 10-50,
т.е. для таких полей невозможно существование наблюдаемых эффектов. Для
волновых торсионных полей с кручением Картана (динамическое кручение) константа
спин-торсионных
взаимодействий теоретически не ограничена. Для торсионных полей с кручением Риччи или Вайценбека нет органический на величину константы взаимодействий, а,
следовательно, и на интенсивность проявления этих полей. Для торсионных полей с
кручением, порождаемых как компонента электромагнитных полей (электроторсионные взаимодействия) константа взаимодействий
имеет порядок 10-3 –10-4.
12. Так как константа электроторсионных взаимодействий (10-3-10-4) чуть меньше
константы электромагнитных взаимодействий (1/137~10-2), то в естественных
условиях такие торсионные воздействия могут вызвать наблюдаемые изменения или
фиксироваться как наблюдаемые сигналы только в тех объектах, в которых есть
неравновесные состояния, ослабляющие электромагнитные связи.
13. Торсионные поля проходят
через природные среды без потерь. Это является естественным фактором, если
учесть, что квантами торсионных полей являются нейтрино. Полезно отметить, что
без связи с торсионными полями советскими физиками более
десяти лет назад было показано, что спиновые возмущения в спиновой среде
распространяются так, что их нельзя экранировать27.
14. Скорость торсионных волн
теоретически равна бесконечности. Сверхсветовые скорости не являются чем-то необычным
для физики. Они присутствовали в теории гравитации Ньютона, они составляют
основы концепции тахионов. Сверхсветовые скорости впервые наблюдались
экспериментально Н.А.Козыревым28, позже подтвержденные двумя другими
коллективами29,30, а на квантовом уровне Цойлингером31.
15. Все тела живой и неживой
природы состоят из атомов, большинство которых обладают ненулевыми атомными или
ядерными классическими спинами. Учитывая, что все тела находятся в магнитном
поле Земли, за счёт магнитных моментов ядер и атомов, которые являются
следствием наличия указанных классических спинов, возникает прецессия, которая
порождает волновое торсионное излучение. Таким образом, все тела обладают
собственными торсионными полями (излучениями).
16. Так как разные тела обладают
разным набором химических элементов, разным набором химических соединений с
разной стереохимией, и разным пространственным распределением в телах этих
атомов и химических соединений, то все тела обладают строго индивидуальными,
характеристическими торсионными полями (излучениями).
Из перечисленных свойств
торсионных полей становится понятно, почему в экспериментах с ДНК фиксируются
пространственные структуры торсионных полей характеристической формы для разных
объектов. Становится ясно, за счёт чего в экспериментах с ДНК наблюдаются
голографические признаки, в частности, нелокальные взаимодействия, а сами даже
дальние взаимодействия реализуются мгновенно.
Литература
Lolle S.J., Victor J.L., Young J.M., Pruitt R.E.
Genome-wide non-mendelian inheritance of extra
genomic information in Arabidopsis. Nature, 434, 505-509
(2005).
Francis Crick. A personal View of Scientific
Discovery. Basic Books, Inc., Publishers,
Biebricher C.K., Eigen M., Luce R. Product analysis of RNA Generated de novo
by Qb Replicase. J.Mol.Biol., 148, 369-390
(1981).
Biebricher C.K., Luce R.,
Template-free generation of RNA species that replicate with bacteriophage
T7 RNA polymerase.
The EMBO Journal, 15, Issue 13, 3458-3465 (1996).
Choudhury G.K., Kejarival
P.C., Chattopadhyay A. The devices for generation of
phantom leaf effect. J.Inst.Eng. (
Gariaev P.P., Junin
A.M. Phantom leaf effect. Myth or reality? Energy 10, 46-52 (1989). In Russian.
Gariaev P.P. Wave based genome. Monograph.
Gariaev P.P. Wave genetic
code. Monograph.
Prangishvili I.V., Gariaev
P.P., G.G.Tertishny, V.V.Maximenko,
A.V.Mologin, E.A.Leonova, E.R.Muldashev. Spectroscopy of radiowave
radiation of localized photons: path to quantum nonlocality
of bioinformation processes. Sensors
and systems, 9 (18), 2-13 (2000).
Gariaev P.P., Birshtein
B.I., Iarochenko A.M, Marcer
P.J., George G. Tertishny G.G., Leonova
E.A., Kaempf U., The DNA-wave biocomputer.
«CASYS» – International Journal of Computing Anticipatory
Systems (ed. D.M.Dubois),
Gariaev P.P., Chudin
V.I., Komissarov G.G., Berezin
A.A., Vasiliev A.A., Hologrphic
Associative Memory of Biological Systems, Proceedings SPIE — The International
Society for Optical Engineering. Optical Memory and Neural
Networks, 1621, 280- 291.
Allison S.A., Sorlie
S.S., Pecora R., Brownian Dynamics Simulations of
Wormlike Chains, Dynamic Light Scattering from a 2311 Base Pair DNA Fragment. Macromolecules, 23, 1110-1118, (1990).
Hagerman, P. J., and B. H. Zimm.,
Zimm B H., Dynamics of polymer molecules in dilute
solution: viscoelasticity, flow birefringence and
dielectric loss, J. Chem. Phys., 24, 269-78, (1956). General
Electric Research Laboratory,
Rouse P E., A theory of the linear viscoelastic properties of dilute solutions of coiling
polymers, J.Chem.Phys., 21, 1272-80, (1953). Tertyshnii G.G., Gariaev P.P., Aksenov V.A., Leonova E.A., Fomchenkov S.V.,
The formalism of endogenous polarization/holographic managing processes in
organisms. Consciousness and a physical reality, 9,
number 4, 44-50 (2004). In Russian.
Prangishvili I.V., Gariaev
P.P., Tertishnii G.G., Maksimenko
V.V., Mologin A.V., Leonova
E.A., Muldashev E.R., Spectroscopy
of radiowave radiations of the localized photons: an
output on quanum nonlocal bioinformation processes. Sensors and Systems, 9 (18), 2-13 (2000).
In Russian.
Заявление о выдаче патента в
Федеральный институт промышленной собственности России от 20.10.2005.
Регистрационный номер 2005132403. Входящий номер 036308.
Tertyshnii G.G., Gariaev P.P., Aksenov V.A., Leonova E.A., Fomchenkov S.V., The formalism of endogenous polarization/holographic managing processes in organisms. Consciousness and
a physical reality, 9, number 4, 44-50 (2004). In Russian.
Prangishvili I.V., Gariaev
P.P., Tertishnii G.G., Maksimenko
V.V., Mologin A.V., Leonova
E.A., Muldashev E.R., Spectroscopy
of radio wave radiations of the localized photons: an output on quanum nonlocal bioinformation processes. Sensors and Systems, 9 (18), 2-13 (2000).
In Russian.
А.Е.Акимов. Эвристическое
обсуждение проблемы поиска новых дальнодействий.
EGS-концепции. МНТЦ ВЕНТ, препринт №7А, Москва, 1992.
Е.Cartan On the Generalization of the Notion
of the Curvature of Riemann and spaces whit Torsion» (Comptes
Rendus Acad. Sci, vol
174, 1922, p.593-595),
Г.И.Шипов. Теория физического
вакуума. М., Наука, 1997, с.450.
М.М.Тернов, В.А.Бордовицын.
О современной интерпретации классической теории спина Я.М.Френкеля. УФН, 1980,
т. 132, вып.2, с.345.
Б.Г.Багров, Б.А.Бордовицын.
Классическая теория спина. Известия ВУЗ, сер. Физика, 1980,Ш, с.67.
Markov M.A. Very Early Universe. Proc. Of the Nuffield Workshop. Eds. Gibbson
G.W., Hawking S.W., Siklov S.T. —
Л.Б.Окунь. Физика элементарных
частиц. Наука, ГР ФМЛ, М., 1988, с.272.
Козырев Н.А., Насонов В.В. О
некоторых свойствах времени, обнаруженных астрономическими наблюдениями. —
Проблема исследования Вселенной, 1980, вып.9, с.76.
А.Е.Акимов, А.Ф.Пугач. К вопросу
о возможности обнаружения торсиопных воли
астрономическими методами. МНТЦ ВЕНТ, М., 1992, препринт №25, с.19.
М.М.Лаврентьев, И.А.Еганова, М.К.Луцет,
С.Ф.Фоминых. О дистанционном воздействии звезд на резистор. Доклады АН СССР, 1990,
т.314, вып.2, с.352.
Dik Bouwmeester, Jian-Wei Pan, Klaus Mattle, Manfred Eibl, Harald Weinfurter & Anton Zeilinger. Experimental quantum teleportation, Nature, v.390,
N11, pp.575-579, December 1997.
Открытое
письмо академику Э. Круглякову — председателю комиссии по борьбе с лженаукой и
фальсификацией научных исследований при Президиуме РАН
Гаряев
П.П.
Случайно, бродя по Интернету,
обнаружил некие «материалы» комиссии по борьбе с лженаукой, подписанные
академиком Э. Кругляковым. В этих «материалах»
фигурирует и мое имя. Цитирую:
«С завидной регулярностью в
газете возникает «биолог» П. Гаряев, труды которого
не находят понимания среди биологов. Впрочем, ясно почему. Речь идет о
совершенно очевидном шарлатанстве. П. Гаряев нашел,
что лазером является сама ДНК человека. Достаточно владеть физикой в объеме
средней школы, чтобы понять, что при поперечном размере ДНК излучение будет дифрагировать в 2 пи стерадиан, так что ни о каком лазере
речи быть не может! Ну а зачем Гаряеву потребовался
лазер? Чтобы переложить наследственные признаки в голограмму. Как ее создать и
как считывать — это уже не обсуждается».
На это мне можно было бы и
промолчать, как вынуждены были молчать и терпеть
биологи-генетики после достославной сессии ВАСХНИЛ 1948 года, когда их громил
«народный академик» Трофим Лысенко. Генетику тогда объявили лженаукой и
«продажной девкой капитализма». Слово «ген» тогда
стало ругательным в генетике. А мы, русские, в то время лидировали в этой
важнейшей области знания. Последствия этого разгрома таковы, что страна
лишились многих блестящих научных умов, и мы отстали, почти безнадежно отстали.
Были преданы анафеме и забыты на десятилетия труды гениальных российских
ученых, таких как А.Г. Гурвич, А.А. Любищев, В.Н. Беклемишев.
А ведь именно они высказали в
20-х — 40-х годах потрясающую по своей прозорливости идею, что гены могут
существовать в форме физических полей, что гены могут образовывать
«гармоническое единство, подобное хору». Гениальное сравнение генетического
кодирования как аналога нотной записи с последующим исполнением этой «генной
музыки» и созданием некого акустического образа будущего организма. Какое
предвидение! Какое опережение своего времени! А.Г. Гурвич создает в те годы
свою знаменитую теперь «Теорию Биологического Поля», где с тем же опережающим
время научным энтузиазмом пытается доказать, что «хромосомы и гены имеют свой полевой
эквивалент».
Но рано, не поняли, вытолкали
А.Г. Гурвича в Германию, а все это направление, связанное и с вещественными, и
с волновыми генетическими функциями, объявили «лженаукой». Если бы тогда не
было гонений на «лженауку», как много бы мы сделали! Как далеко бы ушли вперед!
И вот следствие — наше интеллектуальное отставание в биологии и в генетике.
Запад в этом направлении бурно развивается — открытие двойной спирали ДНК
Уотсоном и Криком в 1953 году и Нобелевская премия им же в 1963 году, затем
последовало открытие генетического кода, и Нобелевские премии посыпались как из
рога изобилия, но не на наших ученых.
В конце 50-х президент США Д.
Эйзенхауэр в одном из своих выступлений бросил презрительно, что СССР отстал от
Запада в молекулярной биологии на 25 лет. Это задело Никиту Хрущева. И тогда, в
1962 году, вышло постановление правительства СССР о мерах по дальнейшему
развитию биологии. Со всех биологических факультетов ВУЗов Союза собрали
наиболее талантливых студентов и направили доучиваться в МГУ на биофак. Я был
среди этих славных ребят и учился у академика Академии Наук, вице-президента
Академии Наук Андрея Николаевича Белозерского на кафедре молекулярной биологии
биологического факультета МГУ, которую он возглавлял.
До сих пор вспоминаю его с
благодарностью. Он поддержал меня, несмотря на мое, обнаружившееся уже тогда,
стремление идти в науке против основного течения. Он предупреждал, оставляя
меня в аспирантуре, что на моем пути будет много людей типа Лысенко и Круглякова. Так что я их не даром сопоставляю. Параллель
очевидная. Не поленился, нашел сайт Круглякова в
Интернете. Даже внешне Лысенко и Кругляков похожи — та же злобная аскетичность и безжалостность в старческих лицах. И
действительно, оба — инквизиторы в науке.
Только Лысенко, худо-бедно,
биолог, правда, временами чудовищно безграмотный. А Кругляков, физик-ядерщик,
но беспардонно залез не в свою область. Решил порассуждать
о биологии и генетике. В добрый путь, г-н Кругляков! Только не мешало бы для
начала в объеме «средней школы» почитать что-нибудь о молекуле ДНК. Однако,
говоря словами покойного академика Фока, «уровень моего оппонента настолько
низок, что не позволяет вести дискуссию». Я имею в виду «знания» Э. Круглякова в биологии и генетике, а также в физико-химии ДНК. Да и газета — не
подходящее место для сугубо научной дискуссии.
Очевидно также, что «знания» Э. Круглякова-обличителя и борца против лженауки
ограничиваются газетными публикациями, что очень четко отметил профессор Л.В.
Лесков, великолепно ответив ему по поводу его «знаний» о торсионных полях,
которые Кругляков отрицает. Как физик Э. Кругляков и здесь, в своей области,
выглядит убого.
Ну а что касается биологии, то
мне даже как-то неудобно выставлять напоказ безграмотность моего оппонента.
Впрочем, он «интересует» меня исключительно как рецидив лысенковщины
от физики с проекцией на биологию, рецидив, который, не дай Бог, снова
затормозит поступательное развитие биологии и генетики в России. Под предлогом
и под шумок борьбы с лженаукой (а с ней надо бороться, и это естественно), на
примере какой-то статьи в газете (а не в научном журнале!) Кругляковы-Лысенки
опять, как слоны в посудной лавке, начнут крушить все подряд, оказывая медвежью
услугу биологии.
Рассмотрим технологию шельмования
в исполнении Круглякова, когда он возмущается мыслью,
что «лазером является ДНК человека». Отвечаю Круглякову
— да, ДНК в хромосомах обладает подобным свойством, она способна к суперлюминесценции. И не только ДНК человека, но и всех
живых организмов. Это предсказано еще А.Г. Гурвичем, он даже пытался обнаружить
излучения хромосом, и с трудом уловил их, и назвал митогенетическими лучами.
А.Г. Гурвич предугадал лазерные свойства хромосом. Гордиться бы этим надо, как
достижением России. Теперь же в этом направлении на Западе работают целые
школы, и весьма успешно.
Например, в Германии
Международный институт биофизики под руководствам Фрица Альберта Поппа
(F.A.Popp), который прямо говорит, что развивает идеи А.Г. Гурвича. А
теперь они работают уже совместно с биофаком МГУ. Но г-н Кругляков об этом,
конечно же, не знает. Тогда пусть посмотрит в Интернете сайт этого Института
или, по крайней мере, спросит у Ф. Поппа, что такое
ДНК-лазер, по электронной почте a0221@rrz.uni-koeln.de.
Да и фундаментальную работу Ф. Поппа и соавторов на эту тему тоже бы надо знать, если
берешься рассуждать на такую сверхсложную тему как волновые кодирующие функции
ДНК. Даю ссылку на эту книгу, заботясь исключительно о расширении кругозора
г-на Круглякова: Popp et al, 1994, Bioelectrodynamics and biocommunications.
Ed. World Scientific.
Мы тоже не стоим на месте в этой
области. Вышли две мои книги:
Гаряев
П.П.,1994, Волновой геном. М. Изд. Общ. Польза. 279с.;
Гаряев
П.П., 1997,Волновой генетический код. М. Издатцентр.
108с.
Особо скажу об экспериментальном
доказательстве возможности работы ДНК как лазера. И не для того, чтобы убеждать
г-на Круглякова в его некомпетентности, толкующего о
каких-то «дифрагированиях в 2 пи стерадиан:» (это в 3-х мерной-то структуре жидких кристаллов ДНК и
хромосом!), а исключительно для осознания существенности самого факта создания
работающей модели суперлюминесценции ДНК. Это
произошло в 1996 году, когда я работал вместе с коллективом блестящих физиков в
одном из крупнейших институтов Российской Академии Наук.
Мы опубликовали в престижном
журнале «Квантовая электроника» наше экспериментальное исследование. Это была
первая в мире работа по лазерной накачке препаратов ДНК и хромосом, работа, в
необходимости которой я лет пять убеждал нашу группу. Конечно, это было трудно.
И убеждать, и экспериментировать. Физики, с которыми я работал, высочайшие
специалисты по лазерам, вообще с трудом воспринимали идею ДНК-лазера, слишком
уж сложна молекула ДНК. Но, в отличие от Круглякова,
они понимали, что в случае успеха открываются огромные перспективы
моделирования лазерных управляющих процессов в организме. Откроются новые
горизонты использования «умных» молекул ДНК как основы для создания
ДНК-компьютеров.
Это огромное направление в
исследованиях, и оно бурно развивается в США, правда, с иных, чисто
вещественных, позиций. Мы же хотели и хотим доказать, что ДНК в составе
будущего ДНК-компьютера может работать и как оптический квантовый генератор. И
это будет именно квантовый ДНК-компьютинг. Так вот,
сумели мы заставить ДНК и хромосомы работать как лазеры и получили генерацию
когерентного света (суперлюминесценцию) генетическими
структурами. Не потрудитесь ли, г-н Кругляков, почитать эту нашу работу,
имеющую и английский вариант? Вот ссылка:
А.М. Агальцов, П.П. Гаряев, В.С. Горелик, И.А. Рахматуллаев, В.А. Щеглов, 1996,
Двухфотонно-возбуждаемая люминесценция в генетических
структурах. Квантовая электроника, v.23, N2,
с.181-184.
Сюда же относится и наши
физико-математические модели, предсказывающие возможность создания лазера на информационных биомакромолекулах:
Гараяев П.П., Юнин
А.М.
Несмотря на успехи
генной инженерии, морфогенез (развитие живого организма из единственной
оплодотворенной клетки) остается загадкой. Мы ничего не можем сказать о том,
что собой представляет и где находится та программа, которая управляет
созданием сложнейшей структуры организма. Развивая концепцию советского ученого
А. Г. Гурвича, полагавшего, что организм строит себя под контролем особого
биополя, авторы предлагают свою гипотезу. (еще раз о тайне умирающих листьев)
В середине 70-х годов советские ученые В. И. Инюшин и В. Г. Адаменко сообщили о неких фантомных эффектах, наблюдавшихся у поврежденных листьев растений при фотографировании их по методу Кирлиан.
Напомним, что в основе его лежит так называемая газоразрядная визуализация, открытая около сорока лет назад советскими учеными С. Д. и В. X. Кирлиан. Метод позволяет исследовать любые, в том числе и живые объекты, которые начинают «искрить», если их расположить между электродами генератора высокочастотных высоковольтных электрических полей. При этом исследуемые объекты должны размещаться так, чтобы между ними и одним из электродов имелся некоторый зазор. Если на одном электроде расположить лист растения, а на другом фотопленку, то искры как бы «вычерчивают» контуры листа и частично его внутреннюю структуру.
Механизм образования и структура разрядов сложны и окончательно не изучены.
Фантомы казались необъяснимыми. В самом деле, если можно получить фотографию целого листа в то время, как у него удалена часть, значит, можно увидеть то, чего нет!
Первая реакция была естественной:
этого не может быть. Фантомы, скорее всего, следствие методических
погрешностей.
|
|
Рис. 1. Обычный фантомный эффект на листе бальзамина. Его
получали и до нас |
Доводы критики сводились к тому,
что, во-первых, результаты, по данным самих же авторов, имели невысокую воспроизводимость (из 100 попыток удачными оказывались не
более 5), во-вторых, используемые высокочастотные генераторы не были
унифицированы, а принципиальные схемы в публикациях не приводились.
Лишь в 1979 г. группа индийских
исследователей во главе с Ж. К. Чоудхари привела не
только убедительные результаты, но и подробную схему генератора, с помощью
которого выявлялись фантомы удаленных частей листа. Однако и этим результатам
не спешили верить — уж очень странно, с точки зрения здравого смысла, выглядело
появление фантомов.
Тем не менее
фантомы вызвали новую вспышку интереса к газоразрядной визуализации. Реальное,
видимое глазом, тем более зафиксированное на фотопленке, изображение того,
чего, казалось бы, уже нет, внушало надежду найти выход из тупика, в котором в
последнее время оказались генетика и эмбриология.
Дело в том, что, несмотря на
успехи генетиков, представления об основных информационных процессах в живой
клетке с момента открытия роли ДНК практически не изменились. Мы знаем, что
клетки по генетической памяти, заключенной в ДНК, нарабатывают основной
материал организма — белки, а они как-то выстраиваются в сложнейшую трехмерную
структуру — живой организм. Вот это «как-то» и остается неизвестным. Мы
практически ничего не можем сказать о том, каким образом оплодотворенная
яйцеклетка «помнит» все детали и размеры развивающегося из нее организма, что
управляет восстановительными процессами в нем, каков механизм получения точной
копии целого организма из единственной клетки предшественника при вегетативном
размножении.
Все попытки вывести трехмерную
структуру организма, исходя из схемы «ген — признак» оказывались
безрезультатными. Задача еще более усложнилась после того, как была открыта так
называемая «эгоистическая ДНК» — часть ДНК (причем большая), которая не
содержит генетической информации, устойчива к
повреждениям и очень подвижна. В недавно вышедшей книге «Эволюция генома» (М.,
«Мир», 1986) фактически содержится признание, что мы не понимаем назначения и
функционирования 95- 98 % генома. Что это — балласт в ключевом
информационном подразделении клетки? Если так, то почему он столь подвижен и не
исчезает в ходе естественного отбора? Чтобы ответить на эти вопросы, нужно
знать, как именно кодируется трехмерная структура организма и где находится
этот код. Вот тут-то и могли пригодиться фантомы. Разумеется, лишь в том
случае, если их существование будет твердо установлено.
|
Рис. 2. Фантомный эффект на листе
березы (а); (б) — схема удаления фрагментов листа. Обращает на себя внимание,
что не все удаленные фрагменты замещены фантомами. Это показывает либо на
частичное разрушение голографических решеток высокочастотным полем, либо на
изменение их линейных размеров |
ЧТО ПОКАЗЫВАЮТ ФАНТОМЫ
Мы решили воспроизвести эксперименты Ж. К. Чоудхари. В Институте химической физики АН СССР была создана установка, принципиально повторяющая индийскую с той лишь разницей, что мы использовали специально разработанный нами водный электрод, описания которого в публикациях не было. Как стало ясно позднее, именно он и обеспечил регистрацию эффектов, ранее не наблюдавшихся.
Типичный, «классический» фантом представлен на рис. 1. Такие получали и до нас. А вот на рис. 2 — необычный фантом на листе березы. По схеме удаления фрагментов видно, что из трех вырезанных кусочков восстанавливается изображение только одного — верхушки листа, имевшей сложный двугорбый профиль. Тот же рисунок имеет и фантом.
На рис. 3 можно наблюдать
характер образования фантома на листе фиалки. По схеме видно, что фантом
возникает только на месте фрагмента 3, удаленного из внутренней части листа.
Однако он исчезает, если удалить фрагмент 4, выводящий вырез в наружную часть
листа. По-видимому, для возникновения фантома существенно расположение выреза.

Рис. 3. Фантомный эффект на листе фиалки зависит от положения
удаленных фрагментов
Совершенно иной тип
фантома (рис. 4) получен на другом листе фиалки. Здесь наружные кусочки листа
не замещаются фантомом, зато кусочки, удаленные из центра, дают фантом, который
представляет собой изображение волосков листа фиалки — то есть конкретных
анатомических структур.

Рис. 4. Фантомный эффект на листе фиалки (а), у которого по схеме
(б) удалены фрагменты. Видны фантомы волосков листа — то есть
конкретные анатомические структуры

Фантом на листе
герани (рис. 5) воспроизводит не только траекторию края отсутствующей части
листа, но и внутреннюю структуру выреза. Любопытно, что этот фантом исчезал,
стоило лишь перевернуть лист на другую сторону. Анализируя «портреты»
несуществующих частей листьев самых разных растений, так и хочется сделать
вывод, что мы видим не что иное, как электрический (или полевой) план достройки
части листа до целого.

Рис. 5. Фантомный эффект на листе герани. Обращает на себя внимание
неравноценность сторон листа в отношении способности образовывать фантом в
конкретных условиях эксперимента
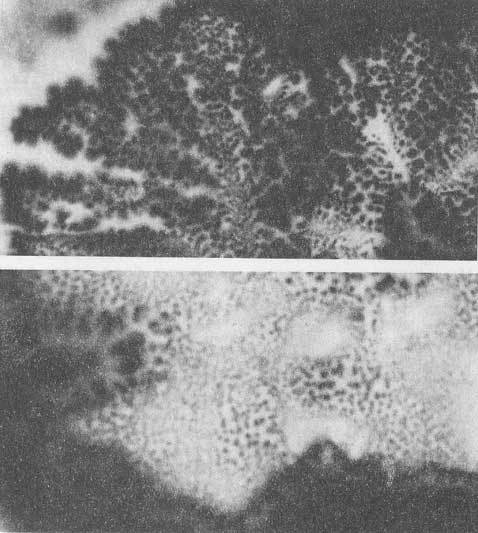
Близкие идеи уже давно
обсуждаются в биологии. Еще в 1944 г. наш соотечественник А. Г. Гурвич выдвинул
концепцию, в соответствии с которой каждая клетка организма генерирует поле,
источником которого являются хромосомы, а объединенные поля клеток дают
направление их развитию в ту или иную ткань, орган, организм.
В те времена концепция биополя носила в общем умозрительный характер и развития не
получила. И вот теперь фантомы как будто подтверждают предвидение А. Г.
Гурвича.
Однако мы не должны забывать, что
все наши фантомы зарегистрированы с помощью газоразрядной визуализации, то есть
как бы искусственно. Если же фантомы действительно представляют собой план
регенерации или развития биосистемы, то они, может быть, проявят себя и без
внешних, а потому чужеродных для организма полей высокой интенсивности.
Поставив серию экспериментов с использованием специальных фотопластинок для ядерных исследований, мы убедились, что целые листья слабо излучают в темноте и засвечивают пластинки даже если между фотослоем и объектом помещены тонкие прокладки слюды. При этом образуются изображения с мельчайшими деталями строения (правда, не в любое время года). Самое же главное то, что в ряде случаев на местах отрезанных частей образовались настоящие фантомы (рис. 6).
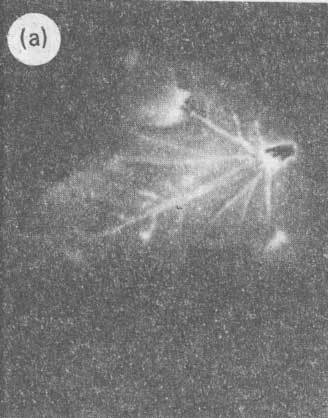
Рис. 6.
Фантомные эффекты на
листьях ясеня, полученные без применения кирлиановской
фотографии:
а) лист экспонировался с 1.10.1987 г. по 12.10.1987 г.;

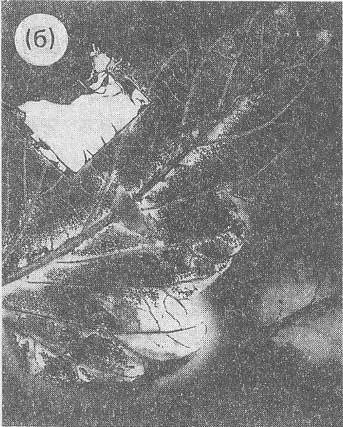
Рис. 6.
Фантомные эффекты на
листьях ясеня, полученные без применения кирлиановской
фотографии:
б) лист экспонировался с 25.07.1987 по 27.07.1987;

Рис. 6.
Фантомные эффекты на
листьях ясеня, полученные без применения кирлиановской
фотографии:
в) увеличенное изображение зоны выреза листа (б) с фантомом
жилкования ткани
Подведем итоги. Фантомы,
содержащие информацию о пространственной структуре листа, существуют в
действительности, и никаких особых воздействий для их визуализации не
требуется. Первоисточником этой информации может быть только хромосомная ДНК в
оставшейся неповрежденной части листа. Значит, и пространственный код
биосистемы в целом размещен там же. Здесь мы вступаем в совершенно
неисследованную область и входим в противоречия с современной генетикой. Мы
допускаем, что хромосомная ДНК содержит не только известный генетический код,
отвечающий за синтез белков, но и сверхгенетический
код фантомов. В этом случае хромосомный набор каждой клетки организма будет
выступать в роли элемента распределенной ассоциативной пространственной (и
временной) памяти о развитии биосистемы и сохранение ее целостности.
О ПАМЯТИ АССОЦИАТИВНОЙ И ГОЛОГРАФИЧЕСКОЙ
Ассоциативная память. Это понятие
сегодня интенсивно разрабатывается в рамках учения о деятельности головного
мозга человека. Основные ее признаки — способность восстанавливать образ или
событие по их фрагментам и отсутствие приуроченности к каким-либо определенным
участкам коры головного мозга. Работает вся кора в целом и каждая ее часть в
отдельности. Этим и объясняются фантомные боли после ампутации, а также
сохранение памяти у людей даже в тех случаях, когда часть мозга разрушена
вследствие травмы или операции.
Ассоциативной памятью обладают и
голограммы. Любой фрагмент голограммы (не меньше определенного размера)
«помнит» образ целого, запечатленного на ней объекта. Избыточность информации и ее помехоустойчивость, присущие ассоциативной памяти,
позволяют сегодня изучать головной мозг как носитель голографической памяти.
Формально в этом смысле и лист растения обладает всеми признаками
голографической памяти. А не является ли фантом изображением голографического сверхгенетического кода? Но тогда возникает вопрос: как
осуществляется запись таких голограмм-кодов и последующее их считывание? Ведь
для этого необходимы лазерные поля и среда, на которой может быть записана
голограмма. Имеется ли такой набор у растений, животных и человека? По
некоторым данным, имеется.
Специалисты ФРГ, развивая идеи А.
Г. Гурвича, показали, что хромосомы растений и животных функционируют как
лазеры с перестраиваемыми длинами волн (от 300 до 800 нм). Что касается среды,
на которой фиксируется голограмма, то роль таковой может выполнять хромосомный
материал, обладающий свойствами жидких кристаллов. Скорее всего, роль носителя биоголограмм выполняет «эгоистическая ДНК», а смысл
подвижности ее участков в том, что они изменяют местную структуру хромосом и
тем самым способствуют или препятствуют «считыванию» фантомов с хромосом
организма. Если это так, тогда снимается один из парадоксов генетики:
становится понятно, почему хромосомы нечувствительны даже к обширным
повреждениям в области «эгоистической ДНК». Это — следствие помехозащищенности биоголограмм.
А существуют ли весомые аргументы
в пользу правильности наших взглядов?
В 1986 г. Абрахам
Зоке из лаборатории физики высоких энергий Стенфордского университета (США) опубликовал результаты
исследований, в которых продемонстрировал, что при облучении жидких кристаллов,
содержащих атомы тяжелых металлов, мягким рентгеновским излучением или
фотоэлектронами вокруг каждого из этих атомов генерируется голографическое
отображение их ближайших окрестностей. Суммируясь, голограммы дают одиночное
отображение уже большей зоны. (У нас в стране подобное теоретическое исследование
проведено В. А. Намиотом в 1988 г.).
Не исключено, что процессы,
описанные этими авторами, вносят вклад и в образование фантомов на поврежденных
листьях и при газоразрядной визуализации и в естественных условиях, поскольку
рентгеновская составляющая присутствует и при газовом разряде, и в качестве
слабого естественного фона Земли.
Развивая мысль об ассоциативной
памяти хромосом, можно предположить, что каждое клеточное ядро и все ядра
клеток организма работают как своеобразный биоголографический
компьютер. Такой компьютер мог бы, во-первых, генерировать объемные образы
(фантомы), по которым строится биосистема, во-вторых, постоянно зондировать
внутреннюю структуру биосистемы для ее самокорректировки,
и, наконец, обрабатывать полевую информацию организма и окружающей среды с
последующим принятием «решения» о генерации того или иного фантома в то или
иное время. В зависимости от этого должны «включаться» те или иные гены для
наработки белков определенного вида.
Ну, а как записываются ДНК-биоголограммы на хромосомах? Ответить на этот вопрос мы
пока не можем. Но почему бы не порассуждать на эту
тему?
В сущности, любая голограмма —
это особый фильтр — преобразователь электромагнитных или звуковых полей. И этот
преобразователь может быть создан, по меньшей мере, двумя путями. Первый —
когда два луча лазерного света пересекают друг друга, причем на пути одного из
них находится интересующий нас объект. Картина взаимодействия лазерных лучей в
виде особого узора (голографической решетки) проецируется на чувствительную
среду (например, желатин) и «запоминается» ею. Если теперь этот узор освещать
лазерным светом, то там, где был когда-то объект, появится точная его копия.
Можно пойти и другим путем. Если заранее знать форму и структуру объекта, узор
голограммы можно начертить с помощью компьютера. В принципе нам ничто не мешает
предположить, что в качестве рисовальщика биоголограмм
мог выступить естественный отбор. Созданные в ходе эволюции узоры в структуре
хромосом преобразовывали внешние поля, которые становились биологически
активными и могли организовывать органические субстраты, клетки и ткани,
которые затем, благодаря организующему голографическому полю, выстраивались в
сложный организм. Таким образом, строение организма попадало в зависимость от
оптических свойств собственных хромосом, то есть от наличия в них
голографических кодов.
ВОДА — УНИВЕРСАЛЬНЫЙ ПЕРЕНОСЧИК БИОИНФОРМАЦИИ
То, что вода является основой
всего живого, известно всем. В воде осуществляются основные биохимические
реакции, перенос жизненно важных веществ, химические коммуникации между
клетками. Казалось бы, что нового можно ждать от воды? Однако именно в этой
тихой заводи, по выражению журналистов, взорвалась бомба. В прошлом году в
журнале «Нейчур» появилась статья 13 авторов из
разных лабораторий мира, которые всерьез взялись исследовать то, что еще
недавно считалось чуть ли не шарлатанством. Рискуя
авторитетом, они занялись Старым гомеопатическим «фокусом», суть которого в
том, что биологически активные вещества, разбавленные водой в чудовищных степенях
(1:10120, то есть в число раз, которое превосходит число атомов во Вселенной),
действуют на человека и даже лечат его чуть ли не
более эффективно, чем известные концентрированные препараты. Срабатывает не что
иное, как «память» воды о веществе — тоже своего рода фантомный эффект.
Любопытно, что у этой сенсации
был предшественник, описавший практически тот же феномен. В 1985 г. немецкий
ученый Ван Д. Кнап опубликовал статью, в которой
демонстрировалось, что биологически активные препараты, будучи бесконечно разбавленными в воде, дают характерное цветное свечение при
газоразрядной визуализации.
В этой связи можно предположить,
что хромосомы, как носители голографических кодов, функционируют также и в
качестве организаторов структуры воды. Благодаря этому «вода» оказывается
способной записывать и хранить голограммы трехмерной структуры организма по
образцу хромосом.
Поддержку этому положению мы
находим в исследованиях Н. А. Бульенкова, который
доказывает возможность образования на молекулах ДНК структур «воды», полностью
повторяющих их строение. Более того, оказалось, что такая «вода» — аналог
различных форм ДНК дает «побочные» ветвления с образованием так называемых
фрактальных структур.
Напомним, что фрактальный рост —
это фундаментальное явление, свойственное как живой, так и неживой природе.
Примерами могут служить ветвление деревьев, рост некоторых кристаллов,
определенные виды электрических разрядов, то есть те процессы, в результате
которых растущая структура в своих частях повторяет самое себя в разных
масштабах.
Для нашей гипотезы наличие водных
реплик ДНК и их фрактальный рост имеет важное значение.
В этом случае голографические решетки способны отображаться в окружающей их
воде и расти. А значит, могут быть «прочитаны» полями с разными длинами волн
без изменения общей картины полевого образа, но с разной степенью подробности.
В результате получается система вложенных один в другой фантомов с разной
плотностью информации. Проще говоря, один фантом даст только контур листа и
грубые детали, а другой — обеспечит картину точного расположения различных
тканей и клеток.
В заключение давайте помечтаем.
Если наша гипотеза окажется верной и голографические коды будут расшифрованы,
мы рано или поздно научимся управлять ими, а значит, сможем «конструировать» и
создавать животных и растения любых форм и свойств. Быть может, разумное
человечество избавится от роковых ныне болезней и вплотную подойдет к решению
проблемы протезирования путем «выращивания» утраченных органов и тканей.
Наконец, овладение биоголографическим компьютером
дает возможность создания его искусственных аналогов,- компьютеров, способных
управлять процессами, сравнимыми по сложности с самой жизнью. Пока же гипотеза
остается гипотезой, и, как всякая другая, она нуждается в проверке и критике.
«Энергия»
Ежемесячный
научно-популярный
иллюстрированный журнал
президиума АН СССР. №10 1989 г
Источник- Академия Тринитаризма - http://www.trinitas.ru/
|
|